| Сегодня 26 июня, пятница |
|
|
|
Какой рейтинг вас больше интересует?
|
Главная /
Каталог блоговCтраница блогера My personal blog./Записи в блоге |
|
My personal blog.
Голосов: 1 Адрес блога: http://abdullin.blogspot.com/ Добавлен: 2010-01-26 23:41:06 блограйдером rusfbm |
|
Очевидное и невероятное в теоретической физике.
2012-04-29 19:03:00 (читать в оригинале)Обухов Ю.А., Захарченко И.И.
22 апреля 2002 года.
Введение.
В 1881 году американским физиком А. Майкельсоном был поставлен исторический эксперимент, призванный выявить скорость Земли относительно эфира. Измерялся фазовый сдвиг световых сигналов в ортогональных оптических линиях интерферометра, оказавшийся равным нулю при любых ориентациях установки.
Впоследствии эксперимент был многократно воспроизведён Майкельсоном совместно с Морли и дал тот же результат. Эксперимент оказал исключительно большое влияние на всё дальнейшее развитие теоретической физики.
Однако следует особо отметить, что интерпретация экспериментов Майкельсона-Морли производилась с позиций классической механики, и в этом проявилась парадоксальность ситуации: выявление эфира и скорости Земли относительно него были произведены с помощью механики, в основу которой заложено равноправие инерциальных систем, исключающее существование абсолютной системы отсчёта и, следовательно, эфира.
Это предопределило ошибочную трактовку опытов, исключив возможность альтернативного объяснения (см. ниже). В итоге был теоретически оформлен отказ от концепции эфира (выразившийся в построении специальной теории относительности - СТО), а принцип равноправия инерциальных систем отсчёта (названный принципом относительности) стал фундаментом современной физики.
Согласно этому принципу, все законы природы одинаковы во всех инерциальных системах отсчёта, то есть уравнения, описывающие физические законы, инварианты относительно преобразований координат и времени из одной системы отсчёта в другую. Этот принцип полная противоположность концепции эфира, для которой характерно неравноправие инерциальных систем и, следовательно, неинвариантность вышеупомянутых уравнений.
Причины всеобщего заблуждения физиков-теоретиков.
Рассмотрим, в чём же заключалась ошибка теоретической физики, утвердившей расчёт опыта Майкельсона-Морли по закону сложения скоростей классической механики.
Научная общественность, воспитанная на классической механике, не имела другого аппарата (иной механики) для оценки результатов опыта, на момент его проведения.
Главная особенность классической механики - это требование мгновенности распространения взаимодействий, то есть законы этой механики справедливы только при условии малости скоростей движения по сравнению со скоростью света. Следовательно, все скорости движений, входящие в Ньютоновскую формулу сложения скоростей (v + с), также должны удовлетворять этому условию.
При расчёте опыта Майкельсона-Морли это условие оказалось выполненным только для скорости Земли (v), второе слагаемое - скорость света (с) - этому условию явно не удовлетворяет. Таким образом, суть ошибки - в нарушении границ применимости механики Галилея-Ньютона. Для расчёта опыта нужна иная механика, отличная от классической и релятивисткой.
Приведём далее главные причины утверждения принципа относительности в физике.
1. Научная общественность, уже имевшая дело с принципом относительности Галилея в механике, легко восприняла то обобщение, каким являлся принцип относительности Эйнштейна.
2. СТО объяснила накопившиеся экспериментальные данные, а в настоящее время её с успехом применяют при проектировании ускорителей элементарных частиц и в экспериментах с релятивистскими частицами.
3. Наконец, сама идея неизменности законов физики при переходе от одной инерциальной системы отсчёта к другой, завораживает своей простотой, изяществом и внутренней непротиворечивостью. Когда физик видит, что преобразования Лоренца могут быть получены из принципа относительности двумя независимыми путями (из постулатов СТО и из условия инвариантности уравнений Максвелла), у него не остаётся никаких сомнений в истинности этого принципа, и вера в принцип относительности становится непоколебимой.
Причины, приводящие к возрождению концепции эфира.
СТО, построенная на основе принципа относительности стала фундаментом современной физики высоких энергий. Тем не менее, есть серьёзные основания для того, чтобы отказаться от принципа относительности:
1. Специальная теория относительности содержит внутреннее противоречие, заключающееся в парадоксе близнецов. Приведём его в следующей модификации. Рассмотрим близнецов-космонавтов, которые стартуют во взаимно противоположных направлениях, каждый на своём корабле, с равной скоростью относительно Земли. Полёт и возвращение космонавтов к Земле предполагается с одинаковой скоростью. Перед стартом близнецы снабжаются двумя комплектами часов. С помощью первых часов обеспечивают полную симметрию движения обоих кораблей (ускорение и торможение кораблей осуществляется по одинаковой программе по этим часам), а вторые (контрольные) запускаются по первым часам в начале инерциальных участков полёта и выключаются в начале участков ускорений. Таким образом, для контрольных часов мы имеем дело только с инерциальными и полностью симметричными ситуациями. К моменту приземления и встречи в исходной точке (точке старта), каждый из братьев в соответствии с СТО видит отставание контрольных часов близнеца, что противоречит здравому смыслу.
2. Существуют современные эксперименты, устанавливающие зависимость скорости света от направления распространения волны и вступающие в противоречие с постулатом СТО об инвариантности скорости света. Серия таких экспериментов была выполнена Стефаном Мариновым (Австрия).
Опыты Маринова выявили направление распространения световой волны, в котором имеет место превышение скорости света "с" на величину 360 ±40 км/с. Несмотря на очевидный результат и простоту идеи эксперимента Маринова, этот опыт до сих пор не воспроизведён ни в одной из лабораторий мира.
Опыт Маринова игнорируется, поскольку существует современные высокоточные интерференционные эксперименты не только с двусторонним распространением (аналоги опытов Майкельсона-Морли), но и с односторонним распространением света (например, интерферометр Маха-Цандера), которые признаются достаточными для опровержения концепции эфира.
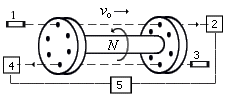
Рис. Схема опыта C. Маринова (развитие опыта Физо
с зубчатым колесом по измерению скорости света).
1 и 3 - лучи лазера, 2 и 4 - фотоэлементы, 5 - детектор тока.
Однако, ошибочность такого мнения заключается в том, что все эти интерференционные опыты, использующие неподвижные оптические линии, принципиально непригодны для обнаружения эфира (математическое доказательство неприемлемости указанных интерференционных методов можно найти в статье на сайте).
Изложенные причины явились основанием для отказа от принципа относительности, что естественным образом приводит к идее возрождения концепции эфира, для которой характерны неравноправие инерциальных систем, с одной стороны, и зависимость скорости света от направления распространения волны с другой.
Альтернативная интерпретация опытов Майкельсона-Морли.
На интерферометре Майкельсона-Морли исследовалось время двустороннего распространения света на отрезке оптической линии длиной L. Был установлен факт независимости (изотропии) этого времени от пространственной ориентации оптической линии: t1+t2=const, t1, t2 - интервалы времени одностороннего распространения света в прямом (от начала отрезка к концу - t1) и обратном (от конца к началу - t2) направлениях.
Сторонники принципа относительности, не имея возможности измерить указанные времена раздельно (ввиду отсутствия соответствующей техники и технологии) и опираясь на принципиально неверный расчёт опыта, трактовали его результат, как равенство времен t1 и t2, отбросив очевидную альтернативную версию: "t1 не равно t2, при условии t1+t2=const".
Если ввести величину, называемую скоростью двустороннего распространения света и определяемую как: с=2L/(t1+t2), то для этой величины (а вовсе не для скорости одностороннего распространения света) из опытов Майкельсона-Морли действительно вытекает изотропия и инвариантность. Такое, казалось бы, незначительное отличие в интерпретации опыта Майкельсона-Морли приводит к диаметрально противоположному результату: к отказу от принципа относительности.
Корректная интерпретация опытов Майкельсона-Морли позволила построить альтернативную механику (СЭТ - "Теория светоносного эфира"). Данная механика, являясь более общей, чем релятивистская, позволяет установить границы применимости последней, а самое главное, приводит к возрождению концепции эфира.
Эфир и космология.
Эксперименты Маринова (1984 г.) выявили значение абсолютной скорости Земли равное 362 ± 40 км/с. Результаты опытов позволяют выдвинуть гипотезу о том, что так называемое реликтовое излучение Вселенной является собственным шумом эфира, поскольку значение скорости, измеренное в опытах, близко к скорости Земли (по отношению к фону реликтового излучения), полученной из астрономических наблюдений.
В этом случае "реликтовое" излучение не является собственно реликтовым, а значит, не служит доказательством происхождения Вселенной по теории Большого взрыва. Другой аргумент сторонников теории Большого взрыва состоит в объяснении красного смещения спектра далеких звёзд эффектом Доплера, вследствие разлёта галактик. Однако существуют альтернативные объяснения.
Например, причинами смещения спектра могут являться: неоднородность эфира - изменение его свойств от центра Вселенной к периферии (в предположении, что наша Галактика находится в центральной области Вселенной), или уменьшение энергии электромагнитной волны вследствие прохождения гигантского расстояния в среде-носителе, при этом поглощённая эфиром доля энергии впоследствии может излучаться в виде шума (предполагается, что процент поглощённой энергии зависит только от пройденного расстояния и не зависит от частоты волны).
Концепция эфира позволяет обосновать более естественный взгляд на Вселенную. Вселенная, как и эфир, являясь вечной, не нуждается в объяснениях своего происхождения. Составляющие её структурные элементы (галактики) непрерывно обновляются: на смену погибающим, старым рождаются новые, молодые.
Иной взгляд на эволюцию Вселенной утвердился в современной астрофизике, объясняющий возникновение Вселенной путём взрыва, связанного с рождением элементарных частиц в результате одного из фазовых переходов в вакууме. Вселенная, в соответствии с теорией Большого взрыва, не вечна, её ожидает гибель либо в результате разлёта галактик ("холодная смерть" - модель расширяющейся Вселенной), либо в результате коллапса ("горячая смерть" - модель осциллирующей Вселенной).
Соответственно, галактики гибнут либо в одиночестве (первая модель), либо коллективно (вторая модель). О том, что теория Большого взрыва является крайне спорной в современной науке, свидетельствуют многочисленные работы учёных - физиков и астрономов.
Так шведский астрофизик, лауреат Нобелевской премии Х. Альфвен говорит: "Эта космологическая теория представляет собой верх абсурда - она утверждает, что вся Вселенная возникла в некий определённый момент подобно взорвавшейся атомной бомбе, имеющей размеры с булавочную головку. Похоже на то, что в теперешней интеллектуальной атмосфере огромным преимуществом космологии "Большого взрыва" служит то, что она является оскорблением здравого смысла: credo, quia absurdum ("верую, ибо это абсурдно")!
Когда учёные сражаются против астрологических бессмыслиц вне стен "храмов науки", неплохо было бы припомнить, что в самих этих стенах подчас культивируется ещё худшая бессмыслица".
От эфира к физическому вакууму - от реального к виртуальному.
Отказ от концепции эфира привёл (с развитием квантовой теории поля) к понятию физического вакуума. Представления о физическом вакууме согласованы с принципом относительности, что предопределило принципиальные отличие понятий эфира и вакуума. Материальный эфир является антиподом физического вакуума.
Действительно, в противоположность эфиру, физический вакуум не является переносчиком электромагнитных волн. Физический вакуум по своей сути является виртуальным объектом, поскольку представляет собой множество всевозможных виртуальных частиц, не поддающихся непосредственной регистрации, при этом выявление скорости тел относительно вакуума принципиально невозможно.
В то же время, эфиру присущи все атрибуты материального объекта: он шумит в радиочастотном диапазоне, относительно эфира можно экспериментально выявить скорость тел и частиц. Так, физика, всегда имевшая дело с измеряемыми величинами, отказавшись от концепции эфира, фактически встала на путь сближения с мистикой, опирающейся на веру в нереальное, виртуальное нечто.
Заключение.
Историческая ошибка, допущенная в начале прошлого века, привела к ситуации, когда принцип относительности приобрёл фундаментальное положение в современной физике, что породило целый ряд заблуждений: отрицание эфира как материальной среды-носителя взаимодействий, создание, в противовес эфиру, концепции виртуальной среды - физического вакуума, построение апокалиптической космологической теории.
Из физической науки стала ускользать физическая картина явлений за математическими выкладками и виртуальными фантомами. Так непоколебимая вера в принцип относительности стала вытеснять логику и здравый смысл: всё противоречащее этому принципу, признаётся заведомо ложным.
Возрождение концепции эфира - это окончательный отказ от принципа относительности и переход к представлению о единстве божественного мира, объединяемого всепроникающей средой-эфиром. Эта среда определяет абсолютную систему отсчёта пространственных координат и времени. Всё, что нас окружает, находится в эфире, более того, рождено из эфира. Фактически эфир - это и есть Абсолютное пространство-время вечной Вселенной.
В социальной и духовной сферах, в которые принцип относительности проник в форме либерализма и политеизма, отказ от относительности морально-нравственных ценностей означает абсолютизацию понятий добра, морали и справедливости.
Справка:
Обухов Юрий Алексеевич, физик.
Захарченко Игорь Иванович, радиофизик, кандидат технических наук.
Обухов Ю.А., Захарченко И.И., "Эфир или физический вакуум", Т-М, 2002, № 10.
Маринов (Marinov Stefan) Стефан (1931 - 15.07.1997).
http://bourabai.narod.ru/marinov/index.htm
Болгарин, гениальный физик профессор С. Маринов в последние годы работал в Австрийском городе Грац. Он описал явление двухкомпонентности магнитного поля, то есть наличия кроме известного векторного магнитного поля еще и скалярной составляющей. На основе своей теории он построил ряд экспериментальных приборов, среди которых "мотор Маринова".
Ему принадлежит открытие способа определения абсолютного движения Земли в эфире. Им определена скорость этого движения - 36030 км/с. Этим своим экспериментом профессор Маринов полностью и окончательно опроверг эйнштейновскую СТО, показав, что скорость света складывается по закону Галилея. Измерив скорость света в одну сторону профессор Маринов показал всю лживость постулатов СТО.
В течение длительного времени, в 1970-1980-х годах, Мариновым была осуществлена целая серия различных экспериментов по измерению однонаправленной скорости света по методикам, принципиально отличающимся от идеи эксперимента Майкельсона - Морли. Результаты экспериментов Маринова с вращающимися дисками свидетельствуют в пользу факта движения Земли в абсолютном пространстве (т.е. относительно неподвижного эфира) со скоростью порядка 360 км/с. Свой фундаментальный эксперимент Маринов осуществил в 1979 году в Брюсселе.
Авторское наименование эксперимента - “coupled mirrors” experiment (эксперимент со связанными зеркалами). Следует признать, что в техническом отношении эксперимент весьма сложен и требует тщательной настройки как механических, так и оптических систем аппаратуры. Основная идея заключается в регистрации изменения скорости прохождения луча света заданного расстояния между двумя синхронно вращающимися зеркалами. При этом Мариновым было установлено, что скорость света, измеренная вдоль выбранного направления на земной поверхности, различна в разное время суток (а значит, - при разной ориентации относительно абсолютного пространства).
По мнению Маринова, “в последнее время, точнее - десятилетия, постулат Эйнштейна о постоянстве скорости света вдоль всех направлений в любых инерциальных системах отсчета приобрел столь устойчивую популярность, что для большей части физиков эта проблема оказалась закрытой, как, скажем, проблема вечного двигателя. Тем не менее, до настоящего времени экспериментального доказательства этого эйнштейновского постулата в пределах первого порядка точности в отношении v/c нет. Исторический эксперимент Майкельсона-Морли, обеспечивающий неприкосновенность догмы о постоянстве скорости света, дает, как известно, точность второго порядка в v/с, но эффекты первого порядка, на самом деле, при этом не могут быть обнаружены. Таким образом, отрицательные результаты опыта Майкельсона - Морли не могут трактоваться как решающее доказательство в пользу концепции постоянства скорости света”.
Обнаруженная анизотропия позволяет выделить направление, совпадающее с направлением движения солнечной системы относительно фонового микроволнового радиоизлучения Вселенной (реликтового фона).
Стефан Маринов явился основателем и директором Института фундаментальной физики в г. Грац (Австрия).
Свои исследования Маринов завершить не успел. 15 июля 1997 г. профессор Маринов погиб, будучи по одной из версий, выброшенным неизвестным из окна университетской библиотеки в центре г. Грац, по другой, согласованной с местными властями - покончил жизнь самоубийством. Странным является поведение полиции, которая не сообщила родственникам Маринова о его смерти, хотя все координаты их имелись в полиции. Его сын - замминистра промышленности Болгарии М. Маринов узнал о гибели отца от его друга только через 2 недели, когда проведение судмедэкспертизы уже проблематично. Преступника не нашли, а дело закрыли, как это было уже не раз в случаях покушения на жизнь изобретателей, опровергавших догматы эйнштейновской теории.
1. Marinov St. Rotating disk experiments / Found. Phys. 1978. V.8. № 1-2, P.136-156.
2. Marinov St. The velocity of light is direction dependent / Czechosl. J. Phys. 1974. B24. № 9. P.965-970.
3. Marinov St. A reliable experiment for the proof of the space-time absoluteness / Phys. Lett. 1975, A54. № 1. P.19-20.
4. Marinov St. Measurement of the Laboratory’s Absolute Velocity, General Relativity and Gravitation, 1980, vol. 12, № 1, р.57-65.
5. Marinov St. Moving platform experiments / Indian J. Phys.1981. B55. № 5. P.403-418.
6. Marinov St. Measurement of the one-way speed of light andthe Earth's absolute velosity / Proc. 2 Marcel Grossman Meet.Gen. Relativity, Trieste, 5-11 july, 1979. Part A. Amsterdam e.a., 1982, P.547-550.
7. Marinov St. Rotating coupled mirrors experiments / Ind. J.Theor. Phys. 1983. V.31. № 2. P.93-96.
Маринов С., «Экспериментальные нарушения принципов относительности, эквивалентности и сохранения энергии» / Физическая мысль России, 1995, № 2, стр.52-77.
http://bourabai.narod.ru/marinov/fmr.htm
veinik.ru
Заговор в науке
2012-04-29 18:52:00 (читать в оригинале)Бурлаков М.П.
1997
"Общественное объединение исследователей и журналистов, известное читателя "Труда" как комиссия "Феномен", создает уникальный информационный банк. В нем собираются любые (даже самые невероятные и фантастические) идеи и открытия, не находящие признания у современной науки. В специальном архиве уже хранятся чертежи "машины времени" и антигравитационного двигателя, расчеты электростанции, действующей на энергии искусственного смерча, описание биотехнологий, позволяющих создавать лекарства с необыкновенными свойствами и многое-многое другое.
Вот лишь несколько "единиц хранения" информационного банка "феномена":
- Специальное покрытие, уменьшающее силы сопротивления движущихся объектов. Если его нанести на крылья авиалайнера, дальность полета увеличится почти на треть. Защищено патентом.
- Уникальный аккумулятор, способный храниться в заряженном состоянии до 10 лет без потери характеристик. Перезаряжается легко и быстро, как затвор у винтовки - простой механической заменой отработанного анода. Защищено патентом.
Всего в банке "Феномена " зарегистрировано уже более 500 ученых, инженеров и изобретателей, работающих в области альтернативной энергетики.
Атомные и гидроэлектростанции дают лишь малую долю в общем производстве электроэнергии. Солнечные, термальные и ветровые установки тоже заведомо не смогут заменить нефть и газ. В такой ситуации, казалось бы, государство должно проявлять повышенный интерес к любым новым идеям, касающимся вопросов производства энергии. Однако, как показал выборочный анализ, проведенный "Феноменом ", ни одна из восьмидесяти взятых наугад (уже запатентованных!) разработок в области энергопроизводства и энергосбережения, не только не внедрена, но даже и не опробована.
Что уж говорить о совершенно новаторских работах!" (Газета "Труд" 19 февраля 1997 г., статья Игоря Царева, "Это просто фантастика", в сокращении. - МБ).
Прочитав в газете "Труд" статью Игоря Царева "Это просто фантастика", я подумал, что наверное, так же, как автор статьи, большинство людей, конечно, испытало два чувства: восхищение по поводу сделанных творческим гением людей изобретений и недоумение по поводу того, что эти изобретения "совершенно никого не интересуют", кроме небольшой группы наивных журналистов-энтузиастов. Я с сожалением подумал, что абсолютно лишен возможности испытать такие же простые и наивные чувства по этому поводу. Прежде всего, я не могу испытать наивное восхищение, потому что знаю: открытий и изобретений в области альтернативной энергетики, и вообще в принципиально новых направлениях отнюдь не 500, а, как минимум, раз в 10 больше, и сделаны они в России. Что же касается удивления по поводу их "невостребованности", то здесь у меня еще меньше возможности для того, чтобы испытать простые и непосредственные чувства, подобно автору этой заметки, поскольку в силу своих профессиональных обязанностей я достаточно долго занимался именно этими вопросами.
Все это можно считать необходимой литературной преамбулой к тому, что изложу далее.
Вторая половина двадцатого века прошла под знаком противостояния двух сверхдержав - СССР и США, распространивших свое влияние на значительной части политической карты мира. По своему накалу, напряжению ресурсов и бескомпромиссности это противостояние имело характер мировой войны; оно и вошло в историю под название "Холодная Война".
Спектр противостояния в холодной войне охватывал, практически, все сферы жизни общества - от пропаганды, создающей противнику образ "империи зла", до напряженной гонки вооружений, изматывающей экономику противника и имеющей целью создать стратегический перевес. Однако, паритет технологий в области вооружения и колоссальная разрушительная сила ракетно-ядерных сил сделали исход прямого вооруженного конфликта неприемлемым для Соединенных Штатов: в случае массированного термоядерного удара территория Северной Америки могла превратиться в радиоактивную пустыню.
В этих условиях ставка была сделана на тайные методы "внутреннего разложения" научно-технического потенциала противника. Успеху этой грязной войны содействовала иллюзорная и лживая идеология, на которой была построена внутренняя политика Советского Союза, отдающая приоритет абстрактным идеям в ущерб национальной безопасности. С учетом этой политики, аналитическими центрами НАТО была разработана концепция подрывной деятельности, имеющей целью ослабить (а в последствии полностью ликвидировать) интеллектуальный потенциал СССР.
В 1991 году автору этой статьи попала в руки копия документа под общим названием "Silver Key", представляющего собой свод инcтрукций для разного рода "фондов" и "научных обществ", разворачивающих свою деятельность в Советском Союзе. В этом весьма любопытном манускрипте имеется раздел, содержащий методы подавления интеллектуального потенциала, и я приведу здесь некоторые "рекомендации" из этого документа, чтобы у читателя не оставалось иллюзий о том, что происходящее в нашей стране есть просто результат некомпетентности чиновников от науки или неудачного курса экономического развития.
С педантичностью, присущей составителям такого рода документов, каждый пункт инструкций снабжен собственным подзаголовком, и мы приводим почти дословный (построчный) перевод некоторых пунктов этого документа:
False aim - ложная Цель, ...Весьма важно навязать противнику ложные ориентиры научных исследований. Для этого можно использовать "рекламу "той или иной научной теории, концепции технологического развития, методики исследований. Определив противнику неверное направление исследований (или заведомо недостижимую Цель этих исследований) можно нейтрализовать работу целых научных коллективов (Лаборатории, КБ, НИИ).
Ложная Цель поставленная перед научным коллективом (или даже перед целой отраслью) является действенным механизмом" омертвления финансовых и материальных средств, выделяемых на научно-технические разработки... Попутно это дискредитирует в глазах начальства конкретный научный коллектив...
Неверная методика исследований приводит к "консервации" перспективной научной идеи и способна отбросить противника в той или иной отрасли на годы (и даже десятилетия)...
Психологической обработке легче всего поддаются руководители научных коллективов, которые, как правило, мало разбираются в конкретной тематике и осуществляют лишь общее (партийное) руководство. Однако, это не лишает их амбиции "крупного ученого", каждое слово которого коллектив должен воспринимать как руководство к действию... Его (руководителя научного коллектива) можно пригласить на международную конференцию (симпозиум), где исподволь внушить необходимое направление исследований... Эффективными являются так же совместные публикации - это привязывает соавторов к определенному научному направлению. Даже если в дальнейшем научная несостоятельность данного направления будет очевидна, "соавтор" все равно будет его отстаивать, используя все (в частности, административные) возможности...
Добавить к этому нечего, можно только проиллюстрировать данную методику в действии на конкретных (крупных) примерах.
Здесь, пожалуй, необходимо привести банальные, но необходимые факты, которые, видимо, каждый человек знает, но в силу их несистемного восприятия, то есть вне контекста всего происходящего, мало задумывается над ними.
А задумываться надо. "...На планете газа осталось на 22 года, меди - на 21 год, свинца - на 21 год, золота - на 9 лет, ртути - на 13 лет, вольфрама - на 2 года". (Данные института материаловедения Академии Наук, газета "Комсомольская правда" за 8 мая 1988 года).
"...Нефти осталось только на 30 лет...". (Данные на 1984 год, - книга "Более чем достаточно?" Издательство "Энергоатомиздат", Москва, 1984 год.)
"...Через 17 лет кончится серебро, через 19 - цинк". (Издательство "Киев", 1990 год).
И так далее, в отношении всех остальных невосполнимых сырьевых ресурсов, совершенно незаменимых для жизнедеятельности современной цивилизации. Здесь мы хотели бы обратить ваше внимание на то, что в действительности основных невосполнимых сырьевых ресурсов на территории современной России залегает до 60% от всех мировых запасов, и приведенные выше цифры уже являются средним арифметическим, то есть наши 60% объединены с оставшимися 40% и разделены на потребности всего населения земного шара, без нашего с вами ведения. Но даже при этом, всех объединенных и разделенных на потребности всего населения земли, невосполнимых ресурсов хватит не более чем на 25-30 лет.
Возвращаясь к прагматическому изложению темы, необходимо сказать следующее.
В условиях истощения традиционных энергоносителей (нефть, уголь, газ) и экологической ненадежности ядерных электростанций, стратегическим направлением научно-технического поиска новых источников энергии считается разработка термоядерных установок. В начале семидесятых годов в этом направлении советская наука достигла значительных результатов, создав уникальные установки, удерживающие раскаленную плазму значительное время в тороидальных магнитных полях. Наша наука подошла вплотную к управляемой термоядерной реакции, значительно опередив западных конкурентов. И тогда была запущена мощная пропагандистская кампания дискредитации самой идеи тороидальных магнитных установок. В научной и околонаучной печати на Западе маститые физики доказывали, что циклические магнитные поля не могут удерживать разогретую до миллионов градусов плазму, приводили выкладки и теоретические соображения. И цель была достигнута! Высокие чиновники от науки поверили зарубежным "авторитетам". Несмотря на явные успехи экспериментов, программа управляемых термоядерных реакций была заморожена и в дальнейшем вообще свернута. Ну, а что же американцы? Через двадцать лет они возобновили исследования по управляемым термоядерным реакциям, которые мы свернули в начале семидесятых годов.
Другой, еще более поразительный пример относится к исследованиям в Космосе. Успехи советской пилотируемой космонавтики ни у кого не вызывают сомнения и особенно это касается создания постоянно функционирующей околоземной обитаемой станции. В этом направлении наша наука обгоняла космического лидера Запада - США - на десятилетия. Увлекшись эффектной с виду, но бесперспективной по сути (в силу целого ряда экономических, технологических и экологических причин) идеей "космических челноков", Соединенные Штаты в первой половине восьмидесятых явно проигрывали космическую гонку Советскому Союзу. И тогда специалисты "психологической войны", используя все рычаги воздействия (вплоть до банального подкупа тех, от кого зависели решения по космическим программам) подтолкнули СССР к принятию своей программы создания "космического челнока", отвлекая материальные и интеллектуальные ресурсы от программ постоянно функционирующих обитаемых орбитальных станций. На свет появился "Буран", но целый ряд перспективных космических программ оказались замороженными.
Теперь Соединенные Штаты пошли по нашему пути, создав проект обитаемой орбитальной станции "Альфа". Но и здесь не обошлось без грандиозного обмана. США предложили России совместный проект станции "Альфа", а когда мы передали им отработанные десятилетиями технологии, Конгресс США предложил исключить Россию из числа участников проекта "Альфа" по причине финансовой несостоятельности. Большую наглость трудно себе вообразить!
Кстати сказать, успех научно-психологической диверсии с "космическими челноками" породил еще более колоссальный блеф шулеров от науки с берегов Потомака - в разработку была запущена басня под названием "Звездные войны" Рональда Рейгана. И советские правители, напуганные компьютерными мультиками, решили сдаться блефующему актеру от политики.
Другой раздел упомянутого выше документа носит название
Our leader - наш лидер. В этом разделе предписывается в каждой научной школе, в каждом научно-исследовательском коллективе, для каждого перспективного направления исследований "создать своего лидера". Под "своим лидером " в этом документе предполагается человек, ориентированный на "западные ценности". Спектр этого понятия достаточно широк: от национальной принадлежности (желательно, чтобы это был, по меньшей мере, нерусский - Е.Т. Гайдар как-то в порыве откровенности сказал, что он не мыслит таланта без примеси семитской крови) до материальной заинтересованности (например, счет в западном банке и вид на жительство, скажем, в США).
Авторы вышеупомянутого документа предлагают разные пути продвижения "своего лидера" - зеленая улица его научным публикациям в западных журналах из этих способов - самый безобидный.
"Свой лидер" - фигура весьма полезная. Он может вовремя посоветовать при выборе направления или методики исследований (важный элемент в организации False aim), может он при случае и остановить не в меру прыткого начинающего научного сотрудника, шепнув кому надо, что исследования, проводимые этим сотрудником, не представляют научной ценности и, вообще - это бред какой-то, или задержать публикацию статьи. Поле деятельности "своего лидера " в науке весьма широко.
Методика "Our leader" давно апробирована мастерами закулисных махинаций и околонаучной возни. Достаточно вспомнить, как автор одной статьи по теории пространства-времени (практически, не содержащей никаких новых результатов и в значительной мере повторяющей более раннюю статью А. Пуанкаре) был возведен на пьедестал "создателя самой революционной теории" и "гения физики двадцатого столетия". Альберт Эйнштейн сполна отплатил своим благодетелям: начинающий физик из Кенигсберга Теодор Калуца прислал в 1919 году Эйнштейну статью о пятимерной теории пространства-времени для представления в "Известия Берлинской академии наук". Публикацию этой статьи "гениальный физик" задержал на три года, сломав кенигсбергскому приват-доценту научную карьеру. Зато впоследствии Эйнштейн сам опубликовал несколько статей по пяти мерной теории, а идеи Калуцы получили интенсивное развитие только в восьмидесятые годы.
Можно не сомневаться, что если бы среди пионеров квантовой теории оказался человек с подходящей фамилией, то усилиями "популяризаторов науки" именно он и возведен был бы в ранг "создателя" второго революционного направления физики двадцатого столетия. Увы, Гейзенберг и Шредингер на эту роль не подходили и квантовая теория осталась без "лидера".
Сейчас академик С.П. Новиков, получивший вид на жительство в Соединенных Штатах, продолжает руководить из-за океана одной из кафедр МГУ и присылает из Америки "свое мнение" кому присуждать, а кому не присуждать государственную премию России.
И еще одна рекомендация в упомянутом выше документе.
Spoilt weapon - испорченное оружие. В этом разделе его пользователям (руководителя "обществ", "фондов", "академий") предлагается сконцентрировать свои усилия на внедрении "рыночных" методов руководства наукой. С неприкрытым цинизмом составители документа утверждают, что русских, не имеющих устойчивых "деловых" навыков, просто захлестнет необузданная стихия рынка. Деньги - испорченное оружие, с которым русские проиграют стратегическую битву за научное лидерство, считают авторы рекомендаций. Деньги помогут на корню скупить научные идеи и технологические разработки, деньги закроют дорогу в большую науку "нежелательным элементам", превращая русскую нацию в отсталую массу мелких торговцев, деньги дадут ключ к обладанию материальными и интеллектуальными ресурсами разоренной страны.
Вообще, "рыночная экономика" - это тот рычаг, при помощи которого аналитики с берегов Атлантики и Средиземного моря мечтают установить новый мировой порядок, идолом которого станет Золотой Телец. Под знаком Золотого Тельца Соединенным Штатам Америки удалось реализовать известный план бывшего шефа ЦРУ Алена Далласа - план разложения изнутри великой державы. "Демократические преобразования" и сепаратистские движения региональной номенклатуры, расшатавшие и подорвавшие институты центральной власти СССР, были оплачены долларами, марками, фунтами.
Первый акт трагедии под названием "новый мировой порядок" завершился подписанием беловежского договора о разделе единого государства на "независимые" республики, который разрезал по живому судьбу русского народа, разорвал единую экономику и разделил мощные вооруженные силы, способные противостоять любому агрессору, на ряд небоеспособных национальных армий, больше напоминающих бандитские формирования.
Ради справедливости, необходимо сказать, что президент России Б.Н. Ельцин был против "беловежского соглашения", но угроза диктатуры коммунистов вынудили его это соглашение подписать, при обязательствах Украинской, Белоруской и Казахстанской сторон по восстановлению единого государства, сразу же после ликвидации коммунистического путча.
Как известно, Кравчук, Шушкевич и Назарбаев обманули Ельцина. Но двоих уже нет на политической арене, а президент России пытается исправить то, что произошло в 1991 году на принципиально новой основе.
Но возвратимся к теме. Президент США не скрывал гордости за победу в "холодной войне", когда заявил, что Америка теперь стала единственной в мире сверхдержавой и должна принять на себя функции по установлению и поддержанию "нового мирового порядка". Этот новый порядок, само собой, должен базироваться на системе ценностей американского образа жизни, где Золотой телец возглавляет пантеон идолов религии стяжательства, насилия и беззастенчивой эксплуатации, прикрываемой демагогией о демократии и общечеловеческих правах.
Однако, натовские стратеги прекрасно понимают, что Россия потенциально остается сверхдержавой и в недалеком будущем, преодолев временные экономические и политические трудности, может снова стать грозным соперником США на пути к мировой гегемонии. В преддверии этого нового (и, очевидно, решающего) противостояния наиболее актуальной задачей Запада является наращивание своего научно-технического потенциала и, самое главное, подавление потенциала своего будущего противника.
Именно поэтому "тайная война" против интеллектуального потенциала России не ослабевает и приобретает все более ожесточенный характер. К традиционным методам (изложенным, например, в упомянутом выше документе) добавляются методы прямого воздействия: проникновение западных специалистов в космические и ядерные программы России, интенсивная перекупка передовых технологий и разработок, массированная шпионская деятельность западных спецслужб под прикрытием различных "фондов" (типа фонда Сороса), интенсивное разрушение системы образования и науки под видом рекомендаций по "оздоровлению экономики" и даже прямое уничтожение наиболее перспективных ученых и специалистов - все это звенья невидимой войны против возрождающейся России.
Описанная нами картина тайной войны в науке была бы неполной, если не упомянуть об одной характерной ее особенности. Основным направлением этой грязной войны, ее главным вектором, является именно подрывная деятельность против интеллектуального потенциала противника, а вовсе не развитие собственного научно-технического потенциала. Иллюстрацией этому может служить создание западными "аналитическими центрами" специальных банков идей и технологий. В таких банках аккумулируются различные перспективные научные разработки, идеи и теории, которые остаются в них невостребованными, без дальнейшего развития и реализации.
Этому есть две основные причины. Во-первых, научная элита Запада зачастую не только не может воспользоваться той или иной теорией или научной идеей, но не способна даже понять внутренний смысл и динамику ее развития. В этом случае идея подвергается обструкции и осмеянию. Пользуясь гигантским пропагандистским аппаратом, не сложно внушить "научной общественности" убежденность в абсурдности той или иной научной теории. Так, по меткому замечанию мудрого русского баснописца: "Как ни полезна вещь, цены не зная ей, невежда про нее, всяк к худу толк свой клонит. А ежели невежда познатней, так он ее еще и гонит...".
Типичным примером здесь может служить "Теория Времени" гениального русского астрофизика Николая Александровича Козырева. Теория эта открывает перспективы принципиально новых источников энергии и способов передачи информации. Однако, со стороны западных астрономов и физиков теория Козырева сразу подверглась яростным атакам и насмешкам, а автор был поставлен в положение изгоя в научном мире. Подобных примеров можно привести десятки.
Такое состояние научной элиты Запада не случайно: она формировалась из "высшего общества" - потомков ростовщиков и торговцев, навязавших цивилизации Запада дух наживы и "ценности свободного рынка". И дух стяжательства умертвил творческую силу познания. Потому что нельзя служить и Богу, и "Маммоне".
Есть и другая, более глубинная причина деструктивного развития западной науки. Что бы не говорили апологеты нового язычества, поклоняющиеся Золотому тельцу, об идеологическом противостоянии Востока и Запада, "тоталитарной империи" и "свободного мира", причиной "холодной войны" был передел жизненного пространства и как следствие, ревизия итогов Второй мировой войны. Обществу непомерного потребления "цивилизованного" Запада необходимы источники сырья, энергоресурсы, дешевая рабочая сила, удаленные от процветающих городов Запада места свалок радиоактивных и токсичных отходов и полигоны для размещения экологически опасных производств. Западная цивилизация превратилась в вампира, высасывающего жизненные соки других народов. В качестве основной жертвы аналитики "свободного мира" видят Россию.
Однако, в конце двадцатого века, глобальное жизненное пространство планеты стремительно сжимается вследствие непомерного расходования "цивилизованными странами" невосполнимых ресурсов Земли. Из этого исторического тупика есть два принципиальных выхода. Первый путь предполагает радикальное сокращение популяции людей и установление глобального контроля за деятельностью всего человечества. Этот путь предлагается Западом и в завуалированной форме озвучен и пропагандируется организациями типа "Римского клуба", "Союза обеспокоенных ученых" и аналогичных "неомасонских" и парамасонских организаций. Эти "клубы" и "союзы" прямо предлагают ввести "планирование семьи", "поставить под контроль (чей ?) расходование мировых (!) ресурсов", внедрить "новую этику человеческих отношений". Они апеллируют к "помощи мирового бизнеса и промышленных лидеров", "правительств развитых стран", "всемирного сообщества ... политиков" (Из воззвания "Союза обеспокоенных ученых"). Одним словом, Запад предлагает установить Новый мировой порядок где, конечно, ведущая роль "правителей" отводится "элите торговцев и ростовщиков" - остальные люди представляют всего лишь человеческий материал для строительства этого "порядка" или подлежащий форсированному сокращению.
Другой выход из ресурсно-энергетического тупика цивилизации предполагает неограниченную экспансию Человечества во Вселенную. Такая экспансия возможна при использовании новейших достижений науки.
Но для этого необходима определенная творческая потенция, а также человеческие, территориальные и природные ресурсы, которыми в необходимом количестве обладает только Россия. Возможности России хотя и являются исключительными, но в то же время они являются потенциальными. А для реализации потенции необходима Воля! Национальная Воля! Воля необходима для того, чтобы обеспечить Независимый от тупиковой Западной Цивилизации путь развития России, мобилизовав для этого имеющийся у нас исключительный потенциал. Для реализации данной цели необходимо осознание этого обществом плюс политическая аккумуляция наиболее волевой, дееспособной его части.
Однако, при таком развитии Человеческой цивилизации вырождающаяся "элита" Запада потеряет свои лидирующие позиции, основанные на тотальной власти торговли и денег.
Эти два пути развития Человечества соответствуют двум состояниям Духа: антагонизм культа стяжательства и свободы творчества. В этом глобальном противостоянии Русской цивилизации присуще именно свобода творчества: "Где Дух Господень - там свобода".
История Человечества уже переживала подобные кризисы. Об этом свидетельствуют древнейшие памятники цивилизации наших предков, разбросанные по огромному пространству Евразии. В более поздние времена, на рубеже нового времени, когда Европа задыхалась, опутанная сетью банкиров и ростовщических контор, а монархи и влиятельные вельможи были кабальными должниками "скромных финансистов", португальский принц Энрике (прозванный Мореплавателем) начал отправлять в Океан каравеллы. Перед изумленной Европой открылся необъятный мир заморских стран, и поток золота разорвал ростовщические узы, опутавшие владетельных князей и монархов. Европа вышла из кризиса "замкнутого жизненного пространства" через внешнюю экспансию и достижения научно-технической мысли (тогда это была география, кораблестроение, навигация, астрономия и картография).
Нынешний ресурсно-энергетический кризис, конечно, несопоставим по глубине и масштабам с прежними кризисами "замкнутого жизненного пространства". Непринципиальный выход из него - в свободном развитии "творческой энергии", а не в рыночной стихии денежных магнатов, пытающихся навязать Человечеству новый мировой порядок менял и ростовщиков.
Справка:
Бурлаков Михаил Петрович (1952 г.р.), доктор физико-математических наук (2000, тема «Клиффордовы структуры на гладких многообразиях»). Окончил Чечено-Ингушский государственный университет (1977), с 1980 года работал в ЧИГУ ассистентом кафедры теоретической механики, старшим преподавателем и доцентом кафедры алгебры и геометрии, кандидат физико-математических наук (1985, тема «Дифференциально-геометрические структуры на гладких многообразиях»). с 1988 года – заведующий кафедрой дифференциальной геометрии и топологии ЧИГУ. В 1991 году во время «Дудаевского путча» уехал из Грозного в Тольятти, где в должности профессора заведовал кафедрой геометрии Тольяттинского педагогического института. В декабре 1993 г. избран в Государственную Думу РФ первого созыва. С 1989 года возглавляет общественную организацию «Центр стратегических исследований», объединяющую специалистов в разных областях науки и технологий.
veinik.ru
Кого будут жечь на костре лженауки?
2012-04-29 18:43:00 (читать в оригинале)Лесков Л.В.
«Российская газета», 30 ноября 1999 г.
Президиум Российской академии наук опубликовал экстраординарный документ - "Не проходите мимо! Научным работникам России, профессорам и преподавателям вузов, учителям школ и техникумов, всем членам Российского интеллектуального сообщества". Это обращение разослано всем возможным адресатам в стране.
За последние годы бурных перемен, происходивших в нашей стране, такого не было ни разу. На столь крайнюю меру Академия наук решилась впервые. "От позиции и действий каждого научного сотрудника, - говорится в этом обращении, - сегодня зависит духовное здоровье нынешнего и будущего поколений". Что же конкретно так обеспокоило президиум РАН?
Это не явная неудача реформ, приведшая к разграблению национального богатства и обнищанию народа. Не превращение могучей мировой державы в слабую ресурсовывозящую страну. Не разгул криминалитета. Не ослабление оборонного могущества державы. И даже не свертывание самой науки, бегство из страны лучших специалистов, угасание научных школ.
Ни об одной из этих действительно насущных проблем не говорится в чрезвычайном обращении президиума РАН. Одно из двух: либо Академия сама, без чьей-либо помощи владеет секретом решения этих проблем, либо считает их не самыми важными для сохранения здоровья нации. Но в любом случае в своем обращении к коллективному разуму нации президиум Академии ставит во главу угла две совсем другие проблемы.
Первая из них - это широкое и беспрепятственное распространение мистических верований - астрологии, шаманства, оккультизма, шабаш колдунов, магов, пророков. Вторая проблема - это торсионные генераторы и другие бессмысленные проекты. Ситуация складывается настолько драматическая, что президиум РАН призывает бросить все силы широких кругов научных работников и педагогов России на борьбу с этим морем бедствий.
Наука в осаде - предостерегает президиум РАН. Мутные волны мистики захлестывают страну, угрожая ее духовному здоровью. А за ними медленно поднимается грозный девятый вал торсионных генераторов...
Что можно сказать об этих проблемах? Прежде всего то, что постановка их в один ряд кажется по меньшей мере странной: о колдунах и прорицателях наслышаны все, а вот о торсионных генераторах из ста человек едва ли хоть что-нибудь знает один. И тем не менее опасность обоих нездоровых явлений президиум РАН считает равномасштабной. Но если, по мнению руководства Академии наук, положение настолько критическое, что для решения этих проблем необходимо мобилизовать все здоровые интеллектуальные силы нации, то независимо от степени нашей осведомленности прислушаться к этому призыву - наш моральный долг.
Отвечая на обращение Академии, считаю необходимым высказать свое мнение по обеим проблемам и предложить их для обсуждения читателям "РГ". Правда, поскольку между обеими проблемами, поднятыми президиумом РАН, совсем немного общего, придется рассмотреть их порознь - сначала первую, потом вторую.
Мифологическое сознание неодолимо.
Мистика есть обязательный элемент двух различных, но родственных систем мировоззрения - мифологической и религиозной. Психологические корни мистических верований уходят в мифологическое сознание. Это хорошо известная истина.
Истоки мифотворчества очень глубоки, они связаны с психологией личности, с процессами становления социального на основе биологического. Миф важен для человека потому, что дает ему ощущение психологической защищенности и смысла бытия. И следовательно, пишет историк культуры Алексей Лосев, миф есть необходимая категория мысли и жизни, в нем нет ничего случайного, ненужного. Для мистически ориентированного человека это безусловно подлинная и максимально конкретная реальность.
Но с другой стороны, миф по определению есть абсолютизация одной идеи, как последней, высшей истины. Отсюда неизбежно возникает фетишизация этой идеи, превращение ее в догмат, не подлежащий ни изменению, ни развитию. А потому переубедить человека, вставшего на позиции мифологического мировоззрения, - задача исключительно трудная.
В силу этих причин бороться с мифологическим сознанием почти бесполезно, само по себе оно практически неодолимо. Главная задача состоит в другом - необходим глубокий научный анализ причин, приведших к не в меру буйному размаху мифотворчества и увлечению мистическими верованиями, а на этой основе построение хорошо аргументированной программы устранения этих причин. Лечить надо не симптомы болезни, а саму болезнь. Других возможностей сохранить духовное здоровье нации не существует.
Причины, о которых идет речь, в общем-то хорошо известны. Если наше обнищавшее население все чаще начинает обращаться за помощью к колдунам и целителям, то это происходит прежде всего не потому, что мифологическое мировоззрение набирает силу. Главная причина в другом - лекарства стали многим не по карману из-за бессовестных действий бизнесменов от фармацевтики. Медицина все больше переходит на платную основу. И бороться тут надо не столько с колдунами, сколько с теми, кто наживает капиталы на людской беде.
В борьбе с дурными проявлениями мифологического сознания не надо ничего изобретать, все методологические подходы тут давно и хорошо известны. Но если ничего этого не происходит, значит, это кому-то нужно. Значит, есть круги, заинтересованные в идеологическом оболванивании населения страны, в том, чтобы, уводя в мифологию, отвлечь его внимание от проблем действительно насущных. Что это за круги, хорошо известно всем, в том числе и президиуму РАН.
Мифология и наука эпохи глобального кризиса.
Наиболее характерная примета мировой истории рубежа XX и XXI веков - нарастающее обострение глобального кризиса. Основную роль при этом играют два фактора - нарушение устойчивости биосферы вследствие губительного воздействия индустриальной цивилизации и неостановимо продолжающийся рост численности народонаселения Земли. Течение кризиса обостряется деятельностью ТНК - транснациональных корпораций, - приводящей к дальнейшему углублению противоречий между Севером и Югом, между миром богатства и миром нищеты.
Решающую роль в развитии этого кризиса по апокалиптическому сценарию может сыграть тысячелетняя традиция создавать новые мифы и неукоснительно проводить их в жизнь. Эта привычка тем скорее способна сыграть губительную роль, что новые мифы приобретают, как правило, псевдонаучную форму.
Один из таких мифов - концепция перехода к устойчивому развитию, принятая на Международном конгрессе ООН в 1992 году в Рио-де-Жанейро. Утопический характер программы отмечали почетный президент Римского клуба А. Кинг и академик Н.Н. Моисеев: программа устойчивого развития ориентирована на решение главным образом экологических проблем, в то время как глобальный кризис носит многомерный и многоплановый характер.
Другой современный миф глобального масштаба - геостратегическая программа, которая популярна в США и смысл которой, по словам одного из ее творцов Збигнева Бжезинского, состоит в том, чтобы "узаконить роль Америки как первой, единственной и последней истинно мировой державы". Нет необходимости напоминать, с какой активностью хозяева Pax Americana приступили к проведению этой программы в жизнь. Провал этих планов неминуем - вопрос только в том, какую цену заплатит человечество, а вместе с ним и народ Соединенных Штатов, за упорное желание осуществить эти несбыточные мечтания.
Под знаком буйных пароксизмов нового мифотворчества развивалась и история России последних полутора десятков лет. Начала движение по этому пути команда М.С. Горбачева, которая капитулировала перед идеологией мондиализма - западной теорией единого глобального мира. Это позволило лидерам Запада использовать перестройку в собственных утилитарных целях. В итоге их совместных усилий погиб Советский Союз, развалился лагерь социализма. В то же время сами лидеры западного мира не пошли ни на какие уступки, ни на какие компромиссы.
Позже уже в рамках России прежний мессианский миф о построении светлого коммунистического будущего скоропалительно попытались заменить другим - о возвращении России в "европейский дом" и ее вхождении в мировое рыночное хозяйство в качестве равноправного партнера.
Российская академия наук в целом как наиболее авторитетная отечественная научная организация своего мнения по этим проблемам так и не высказала. Между тем предложения дать общую научную оценку происходящим событиям прозвучали, например, на юбилейном собрании Академии, посвященном ее 275-летию. И если теперь президиум Академии обращается к интеллектуальной общественности с призывом активно противодействовать мистическим учениям и лженауке, то следовало бы вспомнить, что многие проекты, осуществленные отечественными реформаторами, иначе, как лженаучными, назвать трудно. К сожалению, время заранее предупредить об этом было упущено.
Мистика в зеркале глобального кризиса.
"Источник силы или бессилия общества, - пишет А.И. Солженицын в своей книге "Как нам обустроить Россию", - духовный уровень жизни, а уже потом - уровень промышленности. Одна рыночная экономика и даже всеобщее изобилие не могут быть венцом человечества. Чистота общественных отношений - основней, чем уровень изобилия. Если в нации иссякли духовные силы - никакое наилучшее государственное устройство и никакое промышленное развитие не спасут ее от смерти, с гнилым дуплом дерево не стоит".
Несомненно, широкое распространение всевозможных мистических верований, создание сект вроде сатанистов, чуть ли не повальное увлечение колдунами и псевдопророками наносит немалый ущерб духовному здоровью нации. Но напрасно президиум РАН видит опасность в одной только мистике. Не меньший вред наносит ведущаяся СМИ пропаганда других форм мифов, ориентированных на проповедь крайнего индивидуализма, насилия, сексуальной распущенности, самых низкопробных образцов масс-медиа. Для духовного здоровья нации эта пропаганда намного опаснее деятельности колдунов, астрологов и прорицателей.
Соревнуясь в измышлении как можно более привлекательных мифов, новоявленные претенденты в харизматические лидеры провозглашают: "Я единственный, кто знает, как надо! "Человеку, - пишет по этому поводу французский социолог С. Московичи, - нужны твердая уверенность, неоспоримые истины. Ему нужна целостная картина, имеющая единое основание". И при этом не важно, каким будет это основание - либеральные идеи, национал-патриотическая доктрина или марксистская теория. Современные специалисты в области предвыборных технологий прекрасно усвоили эти истины.
Вносят свой вклад в это виртуальное мифотворчество, включая новые варианты мистических учений, и представители отечественной науки. Академик В.В. Струминский в научном издании "Вестник РАН" опубликовал двенадцать теорем, с помощью которых попытался обосновать тезис о том, что Высший Разум Вселенной крайне неудовлетворен человечеством, сорвавшим замыслы духовной субстанции способствовать снижению энтропии. Чтобы исправить положение, человечеству необходимо безотлагательно "приступить к созданию одухотворенных государств на планете", подавить приобретательские инстинкты, исключить хаос рынка, национализм, шовинизм, все формы эксплуатации человека человеком. Того, кто будет противиться проведению в жизнь этих велений духовной инстанции, предупреждает академик, ждут "кары Всевышнего в земной и неземной жизни".
Доктор философских наук В.А. Кутырев опубликовал несколько статей и книгу, в которых провозглашает: нужен союз философии и религии против свободы научно-технического разума. Другой философ и специалист в области геополитики А.Г. Дугин разработал основы консервативной революции - нового мировоззрения, которому предстоит, по его мнению, сыграть главенствующую роль в грядущей истории России. Приняв православие в качестве идеологической основы этого мировоззрения, России предстоит стать партизаном в тотальном противостоянии с мировой талассократической державой - США. В этой всеохватной войне путь к русскому будущему пролегает через крайнюю форму сопротивления, через переступание границ "искусственных юридических норм", не соответствующих истинным канонам русского права. В этой борьбе русской национальной элите предстоит отстоять государство от "ненаших", которые под знаменем враждебного нам мира Юдаики пытаются поставить его под свой контроль и помешать русскому партизану осуществить свою великую историческую миссию - победить светом истины.
Нет сомнений, подобная мистическая пропаганда, соединенная с приемами научной методологии и ориентированная на пересмотр научного мировоззрения и принятие политических решений, намного опаснее для общества, чем деятельность какой-нибудь провинциальной колдуньи, которая обещает легковерным клиентам сильнейший приворот и отворот, снятие порчи, венца безбрачия, а также ритуалы на бизнес и счастье. Чтобы противостоять подобным выходам мистического мифотворчества на высокий академический и государственный уровень, требуется серьезная мировоззренческая и философская работа.
О лженауке, антинауке и псевдонауке.
Одной из самых опасных разновидностей современного мифотворчества стала лженаука. Определить ее можно, воспользовавшись остроумным афоризмом Станислава Ежи Леца: "Ложь не отличается от правды ничем, кроме того, что не является ею". Лженаука - деятельность, научная по форме, но пустая по содержанию: те, кто занимается ею, сознательно производят на свет мыльные пузыри, выдавая их за научно установленные истины.
Можно говорить о нескольких формах лженаучной деятельности. Первая и наиболее опасная из них - антинаука, которая состоит в возведении лженаучных истин в ранг идеологических доктрин государственного масштаба, а потому неизбежно сопровождается травлей настоящих ученых. Собственно лженаука скромна: максимум, на что она претендует, - это государственное финансирование и реклама СМИ.
Еще более скромные запросы у псевдонауки: она не рассчитывает ни на научную масштабность своих достижений, ни на широкую известность, а ограничивается одним только доступом к государственным источникам финансирования. А потому, как показывает практика, именно она представляет собой наиболее распространенный вид лженауки, который, правда, не приносит большого вреда, если не считать напрасно потраченных государственных средств.
В связи с последним обстоятельством хочется сделать одно замечание. Значительное сокращение финансирования научных исследований привело к тому, что поток малодостоверных, псевдонаучных и лженаучных работ не только не увеличился, но скорее всего уменьшился. Нет никаких статистических данных, которые свидетельствовали бы об обратном. А вот число "псевдосенсационных" сообщений в СМИ резко возросло - это естественное следствие наступившей свободы печати. Авторы обращения президиума РАН, очевидно, спутали два совершенно разных явления. Поэтому, продолжая давать объективную оценку заблуждениям и ошибкам в науке, нет никакой необходимости придавать этой по-прежнему нужной деятельности характер широковещательной общенациональной кампании.
За годы советской власти в нашей стране подобные антинаучные кампании развертывались неоднократно - громили "физический идеализм", "буржуазную лженауку кибернетику", теорию химического резонанса. Особенно масштабным был погром биологической науки, устроенный под руководством "народного академика" Т.Д. Лысенко.
Более свежий пример - организованная вице-президентом РАН академиком
Ю.А. Овчинниковым травля Г.Р. Иваницкого, Ф.Ф. Белоярцева и их коллег, трудившихся над проблемой создания перфторана, - искусственного заменителя крови. Овчинникову удалось использовать для своих целей КГБ, партийных руководителей, журналистов и даже советский суд. Результатом развязанной антинаучной кампании явились трагическое самоубийство Белоярцева, устранение члена-корреспондента РАН Иваницкого с поста директора Института биофизики. На долгое время было задержано продвижение в практику очень нужного медицине препарата, способного спасти жизнь десятков и сотен людей. Волею судьбы среди них оказался и сам академик Овчинников, скончавшийся вскоре после этих событий от рака крови.
Где кончается наука и начинается лженаука?
Работа Роберта Манера - первооткрывателя закона сохранения энергии - была отвергнута его учеными коллегами. Ее отказались публиковать в научных журналах. Затравленный Майер тяжело заболел. Георг Ом был уволен с работы за попытки ввести в науку закон, получивший впоследствии его имя. Жесткие столкновения со своими оппонентами довели до самоубийства одного из самых выдающихся физиков Людвига Больцмана. Нет числа изобретателям, жизнь которых сложилась трагически, но плодами трудов которых мы благополучно пользуемся до сих пор.
В борьбе с лженаукой следует проявлять осторожность. Этому учат нас уроки истории. В сущности, лженаука становится по-настоящему опасной лишь в двух случаях. Во-первых, когда ее пытаются продвинуть на уровень проектов государственного масштаба или, во-вторых, когда без должных на то основании пытаются перестраивать каркас существующего позитивного знания. А полуфантастические публикации на страницах журналов типа "Техника-молодежи" никому не причинят вреда. Напротив, они будут способствовать развитию творческой фантазии пытливых молодых умов и умению давать самостоятельную оценку псевдонаучным идеям.
Занимаясь разоблачениями лженауки, надо помнить, что грань между подлинно научными достижениями и ошибками, которые при желании можно было бы без труда объявить лженаучными, иногда бывает весьма тонкой. В таких случаях надо быть очень внимательным, чтобы с мутной водой не выплеснуть и рациональное зерно. Мне вспоминается одна старая статья в американском физическом журнале, авторы которой яростно критиковали работу немецкого физика Oтто Люммера. Они высмеивали его за то, что, наблюдая под микроскопом поверхность графитового электрода, он утверждал, что при повышении давления окружающего газа происходит плавление графита. Между тем хорошо известно, что графит с ростом температуры не плавится, а сублимирует, испаряется.
Заинтересовавшись этим сообщением, я разыскал в Ленинской библиотеке книгу Люммера "Плавление угля и достижение солнечных температур". Книга была издана в Германии в 1914 году. Между прочим, я оказался первым ее читателем - мне приходилось разрезать страницы.
Конечно, критики были правы - плавления графита Люммер не наблюдал. А вот другой интересный эффект он обнаружил: с повышением до 25 атмосфер давления в камере, где горела электрическая дуга, яркостная температура электродов возрастала до 5300-5600 градусов Цельсия. Когда я рассказал об этом эффекте директору Харьковского института мер и измерительных приборов В.В. Кандыбе, он решил воспользоваться им для создания высокотемпературного эталонного источника света.
Известно немало превосходных научно-популярных работ, посвященных разоблачению лженаучных сенсаций. Среди их авторов были также крупные ученые, как И. Лэнгмюр, Н.Н. Семенов, А.Б. Мигдал, Г.И. Петров, А.С. Китайгородский. А недавно в Новосибирске вышла в свет посвященная той же теме книга академика Э.П. Круглякова "Что же с нами происходит?" В этой книге, говорится в аннотации, академик Кругляков "начал бескомпромиссную борьбу с невежеством, мракобесием, лженауками. В простой и доходчивой форме, живым ясным языком автор разоблачает всевозможных шулеров. Книгу следует прочесть каждому здравомыслящему человеку, кому дорого будущее нашей Родины".
Ничего не скажешь, замысел впечатляет. Тем более что автор книги решением президиума РАН назначен председателем только что созданной комиссии по борьбе с лженаукой и фальсификацией научных исследований.
Но вернемся к вопросу о соотношении науки и лженауки. Здесь имеется еще один очень интересный поворот. Лет двадцать пять назад мне пришлось принять участие в работе по тем временам очень секретной комиссии, которой Правительство поручило разобраться по существу в проекте, предложенном директором Института ядерной физики из Новосибирска академиком А.М. Будкером. Речь шла о разработке нового способа уничтожения боевых порядков баллистических ракет на пассивном участке траектории, т. е. в космосе. Академик Будкер брался за решение этой задачи, если для этого будет выделен миллиард рублей.
Оказалось, что в своих расчетах Андрей Михайлович допустил грубую ошибку. Работа нашей комиссии позволила тем не менее выявить пути, ведущие к решению этой задачи. Одновременно стало ясно: за эту проблему лучше всего не браться - предстояло преодолеть слишком много трудностей, затраты оказались бы колоссальными, а успех отнюдь не гарантирован.
Но дальше произошло вот что. Президент США Р. Рейган выступил с программой "звездных войн", которая дала бы в руки американских военных именно такое абсолютное антиракетное оружие. Наш военно-промышленный комплекс охотно принял вызов. И хотя говорили о "несимметричном ответе", деньги были выделены громадные. А стоило бы своевременно задуматься над другими словами Рейгана: "Мы навяжем русским такую гонку вооружений, что с них свалится нижнее белье". Надо признать, что с этой задачей Рейган справился вполне - победу в "холодной войне" одержали США. И если для победы американские стратеги использовали лженаучное оружие "звездных войн", то они прекрасно знали, что делали.
В своей книге академик Кругляков клеймит главного конструктора НИИ радиоприборостроения Р.Ф. Авраменко, принимавшего участие в этой эпопее. "Самый главный сын лейтенанта Шмидта" - такой титул он ему присваивает. Авраменко, утверждает академик Кругляков, был "отцом-вдохновителем программы чудо оружия". Ошибаетесь, Эдуард Павлович, отцом-вдохновителем одного из реальных вариантов такого оружия был ваш учитель - Андрей Михайлович Будкер.
В экспериментах по программе Авраменко принимали участие известные российские физики, в том числе сотрудники лауреата Нобелевской премии академика А.М. Прохорова. "Комизм ситуации состоял в том, - пишет по этому поводу академик Кругляков, - что они с самого начала понимали бредовость постановки задачи". Хочется спросить вас, уважаемый Эдуард Павлович, если Авраменко, который был убежден, что идет верной дорогой, - сын лейтенанта Шмидта, то как тогда называть этих известных вам физиков, не устыдившихся получать от государства деньги за работу, которую они с самого начала считали бредовой?
Эта совершенно безнравственная и беспринципная ситуация, созданная сотрудниками привилегированного научного института, представляется академику Круглякову всего лишь комичной. А лженауку он предпочитает искать где-нибудь на стороне. Если академическая комиссия по борьбе с лженаукой, возглавляемая академиком Кругляковым, берет на вооружение такой метод, то ее задачи окажутся несложными. Незачем в этом случае посещать лаборатории или читать научные отчеты, достаточно ограничиться чтением газет и разговорами со знакомыми "известными российскими физиками".
Не проходя мимо.
Вернемся теперь к книге академика Круглякова. Полностью разделяя его озабоченность широким размахом лженаучной деятельности и мифотворчества в нашей стране, я хотел бы остановиться главным образом на тех недоработках, которые допустил автор.
Прежде всего вызывает недоумение план книги, выбранный автором. Она представляет собой собрание из двенадцати статей, опубликованных им в разных газетах. Девять из этих статей посвящены основной теме - борьбе с лженаукой. Далее следует рассказ автора о собственной научной биографии, заметка о развитии атомной энергетики после Чернобыля и соображения о курсе проводимых в стране реформ. Такой план построения привел к тому, что практически один и тот же текст повторяется в книге несколько раз, что, конечно, не делает книгу лучше.
Малоудачной представляется и глава, в которой автор рассказывает о своем пути в науку. На фоне критических разборов, содержащихся в предыдущих главах и выдержанных к тому же в весьма раздраженном и саркастическом тоне, данная глава очень напоминает нескромную саморекламу - уверен, что академик Кругляков, известный как серьезный ученый, не хотел этого.
Но все это не главное. Гораздо важнее другое - метод, который академик Кругляков положил в основу своего исследования. Он строит свои заключения почти исключительно по газетным публикациям. Он не встречался с учеными, работу которых критиковал, не посещал их лабораторий, не слушал научных докладов, не читал научных отчетов. Поэтому выполненный им анализ многочисленных заблуждений, которые можно классифицировать как лженаучные, носит в сущности вторичный характер. Это привело к тому, что в книге немало неточностей и просто неверных сведений.
В перечне лженаучных феноменов, которые приводит в своей книге академик Кругляков, главное место занимают торсионные генераторы. На многих страницах книги академик Кругляков детально излагает перипетии своей борьбы по этому поводу с Альбертом Валентиновым - редактором отдела науки "Российской газеты", который имел неосторожность выступить в их защиту. Чтобы переубедить Валентинова, академик Кругляков привлек на помощь своих коллег-академиков В.Л. Гинзбурга и Е.Б. Александрова, но даже и совместными усилиями трем академикам не удалось сломить сопротивление упорствующего журналиста.
Нам остается только посочувствовать академикам и перейти к рассмотрению проблемы торсионных генераторов по существу. Тем более что именно к этой проблеме президиум РАН в своем обращении привлек особое внимание.
Торсионные генераторы и квантово-вакуумные технологии.
Академик Кругляков возмущается прежде всего тем, что на работы по торсионным генераторам затратили 500 миллионов "старых, еще советских" рублей, которые были отняты у нашей нищей армии. Здесь все неверно. Во-первых, в годы, о которых идет речь, наша армия отнюдь не была бедной. Удовлетворялись практически любые запросы военных. Например, когда они заявили, что им необходим космоплан "Буран", на его создание было израсходовано 13 миллиардов рублей, бывших тогда вполне полновесными. А после того, как "Буран" совершил свой первый триумфальный полет в автоматическом режиме, Министерство обороны отказалось от него, заявив, что "Буран" ему не нужен. Именно непосильные для народного хозяйства траты на военные нужды явились одной из главных причин нынешнего тяжелого кризиса.
А вот 500 миллионов для торсионных генераторов никогда не были выделены и выделять их никто не собирался. Суммами такого масштаба распоряжались только ЦК КПСС и Правительство страны. Сведения, которыми пользуется академик Кругляков, - всего-навсего миф, порожденный клеветническими слухами. Возглавляемый А.Б. Акимовым Международный институт теоретической и прикладной физики Российской академии естественных наук из государственных источников финансирования не получает ничего. При желании проверить эти факты не составляло никакого труда.
Следующий вопрос, на который необходимо дать ответ: существуют ли научные основания для создания торсионных генераторов? Вот что думает по этому поводу известный специалист по теоретической физике академик В.Л. Гинзбург, письмо которого академик Кругляков цитирует в своей книге. "Физики знают, что микро- и макромир управляются четырьмя силами. Попытки найти пятую силу безуспешно ведутся последние пятьдесят лет. При этом физики отдают себе отчет, что ищут нечто неимоверно слабое, до сих пор ускользающее от наблюдения". На фоне этих исследований, продолжает далее Гинзбург, вдруг появляются сообщения, что Акимову удалось создать торсионные генераторы, в которых эта сила не только обнаружена, но и работает.
"Безграмотный или фальсифицированный эксперимент может дать любой результат, - пишет по этому поводу Гинзбург. - Справедливость эксперимента подтверждается независимой экспертизой и независимым воспроизведением".
И в самом деле, если торсионное, или спинорное, излучение до сих пор физиками не зарегистрировано, то о каких генераторах этого излучения может идти речь? Чтобы снять сомнения в этом вопросе, обратимся к научной литературе.
В. де Саббата и К. Виваран в журналах "Нуово Чименто" (1989) и "Интернэшнл Джонал оф Теоретикал Физикс" (1990) сообщают о наблюдении сильных спин-торсионных взаимодействий при рассеянии поляризованных протон-протонных пучков и об обнаружении торсионных взаимодействий как пятой силы. В 1976 г. А. Там и В. Хаппер обнаружили, что при прохождении двух поляризованных лазерных пучков через пары атомов натрия эти пучки притягиваются, если они поляризованы одинаково, и отталкиваются при противоположной поляризации. П. Найк и Т. Прадан объяснили этот эффект аксиально-векторным торсионным взаимодействием. Спинорное взаимодействие ядер атомов в квантовой жидкости - поляризованном гелии-3 наблюдали М. Ледюк и Б. Кастэн ("Ля Речерч", 1988). Х. Хайасака и С. Такеучи обнаружили изменение веса вращающихся гироскопов ("Физикал Ревью Леттерс", 1989). Причем эффект наблюдался только в случае правого вращения гироскопов. Этот гироскопический эффект наблюдался и другими авторами (Н.А. Козырев, С.М. и О.С. Поляковы, И. Акира и др.). У. Лале, Д. Фрид наблюдали спиновые эффекты в атомарном водороде и гелии-3, приводящие к резкому изменению свойств газа. Известен образ экспериментальных исследований, проведенных по методологии Этвеша с целью доказать существование пятой силы (Э. Фишман, "Аналс оф Физикс", 1988). Перечень экспериментальных работ, в которых получены убедительные доказательства существования пятого фундаментального взаимодействия, можно продолжить.
Но если академики Гинзбург и Кругляков ошибаются, полагая, что пятая сила до сих пор продолжает ускользать от экспериментаторов, то, быть может, они правы, когда утверждают, что надежных результатов по исследованию торсионных генераторов пока нет? Чтобы снять и эти сомнения, обратимся к результатам экспериментов, проведенных в независимых и авторитетных научных организациях.
В руках у меня протокол по результатам исследования воздействия спинорных излучений на процесс кристаллизации при остывании расплава металла. Протокол утвержден академиком АН СССР вице-президентом АН Украины директором Института проблем материаловедения В.И. Трефиловым. "Проведенные эксперименты показали, - говорится в протоколе, - что при воздействии генератора спинорных излучений на расплав металлов наблюдаются четко выраженные структурные изменения металлов, изменение их физических свойств".
Второй документ - отчет Института физики АН Украины, утвержденный заместителем директора И.А. Солошенко. В отчете подведены итоги исследований влияния излучения генератора торсионных излучений на свойства полупроводниковых и жидкокристаллических структур. Получены однозначные и убедительные доказательства такого влияния.
Третий - отчет Государственного университета телекоммуникаций имени проф. М.А. Бонч-Бруевича (Санкт-Петербург), утвержденный проректором по научной работе С.Л. Галкиным. В работе обнаружен отклик формообразования микрокристалла в сильном электрическом поле на торсионное излучение.
Далее - два отчета Черновицкого государственного университета, утвержденные проректором по научной работе Н.В. Ткачем. Исследовались проблемы создания регистраторов торсионных излучений и влияние этих излучений на объекты с изменяющимися состояниями магнитной подсистемы. Во всех случаях получены положительные результаты.
И еще один отчет - о работе, выполненной в Институте медицинских проблем Севера (Красноярск). Отчет утвержден директором института В.Т. Минчуком. Авторы отчета приходят к следующим выводам: "Установлено, что воздействие торсионного поля на организм человека и взвесь лимфоцитов периферической крови вызывает изменения внутриклеточных обменных процессов, зависящие от индивидуальных особенностей регуляции организма".
Участие МИТПФ РАЕН, руководимого А.Е. Акимовым, в этих и других исследованиях, о которых я для краткости не стал писать, состояло только в изготовлении и поставке экспериментаторам торсионных генераторов. Академики Гинзбург и Кругляков справедливо пишут о необходимости независимого воспроизведения результатов и независимой экспертизе. Но разве многочисленные исследования, проведенные в авторитетных научных учреждениях в разных частях страны, не дают убедительного ответа на этот вопрос? Беда уважаемых академиков заключается, очевидно, в том, что они черпают свою информацию не из научных, а из каких-то иных, им одним ведомых источников.
Столь странные принципы отбора анализируемой информации не позволили ни Круглякову, ни Гинзбургу заметить в пылу полемики с журналистом Валентиновым одно важное обстоятельство: торсионная техника - не единственное направление в области исследования квантово-вакуумных технологий. Эти исследования активно развертываются в последнее время как в нашей стране, так и за рубежом. Профессор Л.Г. Сапогин разработал унитарную квантовую теорию, на основании которой предложил методы получения энергии из квантового, или физического, вакуума. Работы Сапогина опубликованы в зарубежных и отечественных научных журналах. Еще один вариант теории квантового вакуума разработан в серии работ члена-корреспондента Академии наук СССР Ю.Н. Бабаева и Ю.А. Баурова, опубликованных в докладах Академии наук, препринтах ИЯИ АН и в других изданиях. После кончины Бабаева Бауровым проведены многочисленные эксперименты, подтверждающие справедливость теории. Результаты исследований Баурова обобщены в монографии "Структура физического пространства и новый способ получения энергии".
Оценивая практическое значение работы Баурова, академик Н.А. Анфимов пишет: "Предсказывается существование нового взаимодействия в природе... Именно это взаимодействие и может явиться источником силы нового типа, приложенной к космическому аппарату". Речь, таким образом, идет о создании принципиально новых высокоэффективных и экологически чистых космических транспортных систем. Соответствующие эксперименты продолжают в ЦНИИ машиностроения.
Возвращаясь к проблеме торсионных технологий, мне хочется привести большую цитату из статьи, опубликованной в американском журнале "Нью Энерджи Ньюс", где дается оц
Мотор Говарда Джонса как подтверждение теории Эфира
2012-04-29 18:30:00 (читать в оригинале)Магнитный мотор Говарда Джонса является одной из немногих удачных попыток создать практически «вечный двигатель», ибо до сих пор природа сил, которые обеспечивают вращение ротора этого двигателя вокруг статора, так и не получила научного объяснения. То объяснение, которое попытался сделать сам изобретатель непосредственно в патенте, ничего в принципе не объясняет, так как больше направлено на то, чтобы уйти от обвинения в создании «вечного двигателя», потому что Говард Джонс вводит в оборот и призывает на помощь феномен непарных электронов, который, якобы, наблюдается в магнитных материалах. И что именно непарные электроны поставляют энергию этому мотору.
Между тем не все так просто. Ибо даже и такое объяснение идет в разрез с положениями официальной науки. Поэтому мотор Говарда Джонса работает, по мнению официальной, науки с непонятным для них нарушением закона сохранения энергии. И официальная наука как всегда поступает «мудро» и просто делает вид, что такого мотора не существует, или он работает только у самого Говарда Джонса, который знает секрет, с которым он не хочет делиться. Между тем информацию о своем моторе Говард Джонс предоставил для всех желающих с подробным описанием всех деталей этого мотора, и каждый с головой на плечах и некривыми руками может собрать такой мотор.
Благодаря Dragons’Lord’у у нас имеется возможность познакомиться с патентом US 4151431, который получил на свой мотор Говард Джонс. Качество перевода высокое, но думаю, что переводчик помучился с так называемыми непарными электронами, о которых даже в официальных учебниках по физике, по крайней мере, российских, ничего не написано. Не согласен я также и с снисходительным тоном Dragons’Lord’а, что мотор Говарда Джонса – это игрушка, нарушающая закон сохранения энергии. Игрушка, основанная на грамотном управлении эфирных потоков, - это уже не игрушка, а неисчерпаемый источник (генератор) энергии, получаемый из среды, возможности которой мы еще даже не представляем в полной мере. Или иногда получаем представление о ней, когда попадаем в авто или авиакатастрофу, когда на наши дома падает ракета или когда кто-то кому-то разбивает нос. И если кто-то думает, что эфиром тут не пахнет, сильно ошибается.
Ниже зеленым текстом представлен материал по патенту US 4151431. Надеюсь, что читатель осилит этот материал, чтобы в последующем понять, что Говард Джонс совершил такой же подвиг, какой совершил Тесла, создав многофазный мотор переменного тока. Ибо оба типа моторов вращает магнитное поле. Только у Тесла вращающееся магнитное поле задается катушками статора, а уже поле увлекает за собой ротор. А в моторе Говарда Джонса создается «искривленное» должным образом магнитное поле изначально за счет взаимодействия магнитов ротора (арматуры) и статора, и изначально оно жёстко связано с магнитами ротора (арматуры), поэтому искривление «магнитного» поля ротора вращает ротор и вращается вместе с ним. В самом патенте понятия ротор нет, но есть арматура, которая вращается вокруг неподвижного статора. И если понимать принцип «деформации» магнитного поля, который сделал возможным вращение арматуры в моторе Говарда Джонса, то тогда станет возможным мастерить магнитные моторы, основанные на этом принципе, самых разных конструкций. И надо еще понимать, что магнитное поле – это специфическая форма эфирных вихрей, в которых перепады эфирного давления по форме похожи на замкнутый сам на себя винт Архимеда, который вращается по часовой стрелке, если смотреть по его ходу от северного полюса к южному снаружи магнита и от южного полюса к северному внутри магнита. Но более подробно это мы обсудим после того, как познакомимся с патентом.
Патент US 4151431 Мотор Говарда Джонсона
Надеюсь, что читатель понял суть патента Говарда Джонса. Но самое главное, заметил, что магниты арматуры (ротора) «отталкиваются» от магнитного поля статора и двигаются в направлении от южного полюса к северному, при условии, что к ним статорные магниты повернуты северным полюсом. Он отметил также, что изменение полярности только статорных магнитов или магнитов ротора (арматуры) приводит к изменению направления вращения. Почему это так, мы постараемся понять исходя из эфирных представлений. Хотя утверждать, что магниты арматуры отталкиваются от магнитного поля не совсем правильно. Правильнее считать, что за счет взаимодействия магнитных полей магнитов статора и арматуры в области северного полюса магнита арматуры эфирное давление (напряженность магнитного поля) уменьшается, а в области южного полюса магнита арматуры эфирное давление (напряженность магнитного поля) увеличивается, а это приводит к тому, что под действием перепада эфирного давления магнит арматуры начинает двигаться, увлекая за собой и арматуру. Между прочим на таком принципе могли бы летать магнитолеты в магнитном поле Земли или магнитном поле Солнца, если бы удалось бы создать магниты или электромагниты с достаточно высокими показателями создаваемой в них напряженности магнитного поля.
Вся Вселенная, окружающая любого человека (наблюдателя), да и сам человек – это эфир в самых разных фазовых состояниях и проявлениях. Мир – это эфир, находящийся под высоким давлением и в постоянном движении. Мир это потоки и вихри эфира. Мир – это безбрежные пространства между галактиками, заполненные твердым эфиром, частицы которого совершают колебания наподобие тех, что совершают атомы в кристаллах вещества. Каждая галактика, в том числе наша Галактика (Млечный Путь или Молочный след небесной коровы), это эфир, преимущественно находящийся в жидком состоянии, в котором в качестве своеобразных щепок (пены, пара) плавают скопления вещества – звезды, планеты и их спутники, астероиды, межзвездная пыль и газовые скопления, преимущественно водорода. Можно даже считать вещество своеобразной пеной жидкого эфира, но пеной особой, например, как пенопласт, который будучи пеной, имеет высокую жесткость благодаря ячеистой структуре и особенности технологии изготовления. Кроме трех основных фазовых состояний эфир может находиться и в промежуточных состояниях, в виде своеобразного холодца как промежуточной формы между кристаллической и жидкой формами, и в виде безструктурного нестабильного пара, готового к кристаллизации, превращению в жидкость или вещество. Все зависит от состояния окружающего эфира.
В кристаллическом эфире практически нет потоков и вихрей, но есть колебания между частицами эфира и волны, распространяющиеся сквозь него от источников, которые являются процессами в галактиках, звездах, планетах и т.д. Жидкий эфир и его пена (вещество) могут свободно двигаться сквозь кристаллический эфир, так как между его частицами практически нет трения, а сами частицы находятся в постоянных колебаниях, благодаря чему кристаллический эфир ведет себя как зыбучий песок. Кроме того, внедрение в кристаллический эфир жидкого эфира или вещества приводит к своеобразному оплавлению кристаллического эфира вокруг потока жидкого эфира и тем самым создается некий поверхностный слой, в котором перемешаны жидкий эфир и частицы кристаллического эфира (эфирная эмульсия), и этот слой выполняет роль своеобразного подшипника, опираясь на который жидкий эфир проникает в толщу эфира кристаллического, не встречая практически никакого сопротивления.
Жидкий эфир ведет себя практически как идеальная жидкость, но с очень высокой плотностью, и только при столкновении потоков жидкого эфира и его пены (вещества) имеет место незначительная вязкость, которая позволяет веществу вращаться и перемещаться потоками в жидком эфире с одной стороны, а также за счет этой вязкости вещество может вовлекать жидкий эфир во вращение, т.е. вещество при взаимодействии с жидким эфиром способно из жидкого эфира порождать вещество. То есть, та Вселенная, которую мы можем наблюдать, представляет собой тоже эмульсию, но эмульсию из жидкости и пара (газа). Но и жидкий эфир может воздействовать на вещество так, что вещество распадается и пена (пар) жидкого эфира вновь становится жидким эфиром. Именно вращение жидкого эфира порождает такие феномены как гравитационное, электростатическое и магнитное поле. Но об этом чуть позже.
Эфирная пена (эфирный пенопласт) – это совокупность эфирных вихрей, которые уже за счет их высокой энергии вращения приводят к тому, что внутри эфирных вихрей, да и сами вихри – это эфирный пар. Так как скорость вращения стенок вихрей эфирного пара очень высока, то стенки этих вихрей очень жестки по сравнению с «жесткостью» жидкого и твердого эфира. Благодаря этому эфирные конструкции вещества при незначительном трении о жидкий эфир могут существовать длительное время от секунд до миллиардов лет. В зависимости от того как закручены вихри в пенопласте (пене, паре) эфира, вещество тоже может существовать в твердом, жидком и газообразном состоянии. И если о многообразии, например, жидких форм эфира мы можем только догадываться, то о многообразии атомов, молекул, и т.д. мы уже прекрасно знаем, потому что для взаимодействия с этой формой материи у нас имеются развитые органы чувств или мы уже успели создать обилие приспособлений – микроскоп, телескоп, электронный микроскоп, электромагнитные, электростатические и магнитные датчики, дозиметр для измерения опасных излучений, химические тестовые наборы и многое другое.
Сверхбыстрые, с одной стороны, и сверхмелкие, с другой стороны, вихри эфира имеют склонность взаимодействовать между собой, что и ведет к тому, что из атомов создаются молекулы, из молекул создаются уже более крупные образования, такие как горы, океаны. Скопление вещества разного рода порождает планеты, звезды и галактики. Но в любом случае вихри, из которых так или иначе, строятся вещественные композиты, окружены жидким эфиром, который играет роль среды, в которой протекают все известные нам материальные проявления, жидкий эфир выполняет роль смазки для вещества, создает границу между скоплениями вещества, служит средой, внутри которой вещественные образования обмениваются между собой веществом, энергией и информацией. Жидкий эфир сам может создавать мощные потоки, в которые он вовлекает скопления и потоки вещества. Именно вихрь жидкого эфира образует саму Галактику, именно жидкий эфир, по крайней мере, для нас, будучи под высоким давлением, благодаря своей способности взаимодействовать с веществом, является источником энергии. И той средой, в которой «растворяется», конденсируется вещество, после того, как оно израсходует энергию вращения своих элементарных вихрей.
Несколько предположений о природе давления, под которым находится, по крайней мере, жидкий эфир. Это может, во-первых, быть поверхностное натяжение той капли жидкого эфира, в которой мы существуем. Эта капля имеет огромные размеры, сотни и миллионы световых лет. Второй источник давления – это само вещество, которое является для жидкого эфира паром. Как известно, при переходе жидкости в пар объем занимаемый жидкостью увеличивается примерно в 1000 раз. И если хотя бы 0,1% жидкого эфира превратится в пар (пену, пенопласт), то объем такого вещества будет сопоставим с оставшимся жидким эфиром. А в целом объем смеси жидкого эфира и его пара увеличится в 2 раза. Или в 2 раза увеличится давление в том объеме, который занимал жидкий эфир до превращения его незначительной части (0,1%) в пар. А с учетом несжимаемости жидкого эфира повышение давления может быть гораздо выше. И, скорее всего, эфир в виде вещества существует в гораздо меньшем «объеме», не более 0,001%. Тогда вещество будет занимать уже не 50% от объема, а всего 1%. Это уже ближе к реальным цифрам, хотя более правильное соотношение между объемом вещества и жидким эфиром можно оценить, сравнив объем звезд с объемом того пространства, в котором эти звезды образуют нашу Галактику. Нужно еще учесть, что и внутри вещества часть объема занята жидким эфиром, хотя доля его снижается от газов до тяжёлых металлов.
Видимо с этим можно связать все рассуждения горе теоретиков о темной материи и энергии. Вещество мы видим, можем его измерить и т.д. А вот жидкий, а тем более кристаллический эфир мы напрямую воспринимать, кроме некоторых уникумов, не можем. А ведь 99,9% и более материи, которая может породить всё, что угодно, это уже очень серьёзно. И пора прекратить тратить народные деньги и сочинять никому не нужные диссертации, чтобы обосновать СТО и ОТО Эйнштейна или найти несуществующую частицу Хигса. Иначе мы так и не поймем, почему в один прекрасный день волна или вихрь эфирного цунами разрушит наш прекрасный мир. Слепцы, не видящие 99,9% материи, а тем более не признающие её существование, не могут быть нашими поводырями.
С эфирных позиций легко объяснить такой феномен как взрывы сверхновых звезд. Это пока только предположение, но допускаю, что после того, как вещество звезды израсходует свой энергетический ресурс, когда все вихри в звезде начнут рассыпаться и превращаться в жидкий эфир, то это будет равноценно схлопывания кавитационного пузырька. Но в данном случае таким кавитационным пузырьком будет звезда, в котором произойдет конденсация эфирного пара в жидкий эфир. Объем звезды сразу уменьшится в 1000 раз. И если скорость схлопывания будет высокой, то звезда, схлопнувшись, породит резкое падение давления эфира в объеме и центре уже погибшей звезды. Окружающий звезду жидкий эфир ринется туда, где эта звезда только что была. И произойдет чудовищной силы эфиродинамический удар, мощь которого, скорее всего, будет способна породить не только новую звезду, но и сформировать вещество для новой галактики. Ибо по закону интерференции в самом центре бывшей звезды давление ударных волн, направленных со всех сторон будет суммироваться, а вот энергия, которая породит ответную реакцию, будет уже пропорциональна квадрату суммарной амплитуды итоговой ударной волны. И под действие этой мощной ответной ударной волны жидкого эфира, вновь созданное за счет завихренности вовлеченного в процесс эфира вещество будет разлетаться по всем направлениям, пока под действием трения с потоками окружающего жидкого эфира и массами кристаллического эфира не начнется сворачивание исходящих эфирных потоков и потоков вещества вместе со скоплениями вещества в новую звездную систему или молодую галактику, в новый достаточно большой вихрь. Хотя может не повезти и тогда образованное вновь вещество разберут на строительные материалы ближайшие звездные системы и галактики. Так что те, кто верит в теорию Большого Взрыва отчасти правы, когда моделируют ситуацию, но только уже после взрыва, хотя они не понимают, что Большой Взрыв был не один. И прежде всего не понимают как эти взрывы и за счет чего происходят. Для Вселенной взрыв сверхновой звезды равноценен разрыву фолликула в яичнике женщины, выпускающей на волю яйцеклетку с целью породить, если повезет забеременеть, новую жизнь, ибо взрыв сверхновой звезды с высокой долей вероятности приведет к созданию новой звездной системы, на которой, если повезет, может сформироваться жизнь и разум. А если повезет, то энергии взрыва сверхновой может оказаться достаточно, чтобы из окружающего эфира испечь новую и красивую галактику.
Раз уж пошла речь о том, что многое в этом мире совсем не то, что нам кажется, то нельзя обойти такой феномен как масса и инерция. По моим представлениям, если измерять массу по наличию эфира в занимаемом теле или эфире пространства, то самым эфиротяжелым будет кристаллический эфир, затем эфир жидкий. Хотя не исключено, что самым тяжелым может оказаться жидкий эфир, подобно тому, как лед оказывается легче воды. Но самым легким в эфирных единицах будет вещество, так как вещество – это пары эфира, хоть и закрученные в вихри, но пары, пена и не больше.
Но для наших органов чувств и измерительных приборов, настроенных на наши органы чувств и в соответствии с нашими наивными представлениями о Вселенной, получается наоборот – вещество тяжелее эфира. А среди вещества металлы тяжелее газов, хотя с эфирных позиций газ – это слегка завихренный жидкий эфир с небольшим количеством вихрей, а металл – это пространство, практически полностью заполненное огромным количеством вихрей газообразного эфира, а значит практически не содержащее жидкого эфира. Почему же так нам кажется?
А кажется нам так потому, что мы привыкли доверять свои органам чувств, которые указывают нам, что перемещение металла с ускорением в эфирной среде будет вызывать более заметное сопротивление со стороны жидкого и кристаллического эфира, чем перемещение такого же объема газа, который практически жидким эфиром и заполнен. Поэтому один и тот же объем металла кажется нам более тяжелым, чем объем газа. Это же касается и веса. Ибо один и тот же объем металла будет прижиматься к Земле сильней, чем такой же объем газа. Ибо через газ эфир свободно проходит, а через переплетение вихрей собственной пены жидкий эфир вынужден проходить с огромным сопротивлением. Отсюда и разница в весе в «поле тяжести» Земли металла и газа, занимаемых одинаковые объемы.
Поэтому с учетом этих факторов, приведших к нынешним ложным представлениям, массу вещественных тел, следует вычислять как произведение объема вещества на разность плотностей эфира вне и внутри этого объема вещества. Поэтому придется будущим исследователям серьезно поломать голову как над определением того, что является плотностью эфира, так и над методами его измерения в кристаллическом и жидком эфире, а также в различных вариантах вещества, эфирной пене. Если следовать формуле, то такого параметра как масса в современном понимании у кристаллического или жидкого эфира нет, ибо ускорение при столкновении вещества с эфиром возникает у вещественных тел, а вот как ведет себя сам эфир мы просто не знаем, ибо методов регистрации того, что происходит в эфире, кроме некоторых случаев, у нас пока нет. Или придется считать массу любого объема кристаллического и жидкого эфира равным нулю, что позволит объяснить многие странности в нашем мире. Короче, физика нуждается в эфирной революции, которая по последствиям окажется более значимой, чем революция, которую совершил в астрономии Никола Коперник.
Вернёмся к так называемым гравитационному, магнитному и электростатическому полям. Все это особой формы потоки в жидком эфире вне вещественных тел или жидкой фазе эфира внутри вещественных тел.
Гравитационное взаимодействие между вещественными телами станет понятным, если представить себе эфирные потоки и вихри, прежде всего жидкого эфира, из которых, по сути, образуются галактики или звездные системы. Наблюдателю, например, сидящему на Земле, кажется, что Солнце притягивает Землю, а Земля Солнце. И даже формулу для «притяжения» между Солнцем и Землей можно создать, которая всем известна. Но это иллюзия. В действительности и Солнце и Земля являются частями гигантского эфирного вихря, точнее пинпонговыми шариками в потоках вихря жидкого эфира, диаметр которого составляет несколько десятков астрономических единиц, в котором наряду с жидким эфиром отмечается высокая доля эфира парообразного (вещества), закрученного в свои вихри. И законы вращения такого вихря таковы, что в этом потоке присутствующее в нём вещество в виде Солнца, планет, астероидов и комет потоком жидкого эфира прижимаются друг к другу. То есть дело обстоит так, что вещественные образования «придавливаются» жидким эфиром друг к другу, поэтому причина «тяготения» проявляется во времени раньше того движения, которое и характеризует сам факт «притяжения». Поэтому и создается впечатление, что скорость гравитационного взаимодействия бесконечно велика. Ибо, какая может быть скорость взаимодействия между вещественными телами, если это взаимодействие по сути дела самими телами и не регулируется, не определяется и не управляется. Любая планета плывет в пространстве и описывает свои эллипсы по воле волн жидкого эфира и не более. Ну, плывет себе цепка в бурном вращающемся потоке, и пусть думает, что это она притягивается к центру этого вихря. Так оно как-то спокойнее, а то придется искать объяснение в некой Божественной силе, эфир-то мы не видим или не признаем.
О электростатике мы говорим, когда наблюдаем взаимодействие так называемых зарядов. Сам феномен проявляется при трении, например, стекла и шелка, янтаря и шерсти. Это приводит к тому, что потертое стекло или потертый янтарь начинают притягивать к себе заряженные тела иной полярности, либо индуцируют создание противоположных зарядов на незаряженных телах и притягивают их. С эфирных позиций дело обстоит так. Примем, что элементарный заряд, это вихрь. И если атом закручен по часовой стрелке, то это положительный заряд, если против часовой, то отрицательный. В незаряженном теле атомы закручены в вихри, половина которых вращается в одну сторону, а половина в другую, вихри, закрученные в разные стороны образуют спарки, которые в нормальных условиях как бы автоматически и обеспечивают то, что заряд тела нейтральный. Но стоит тело потереть, как часть вихрей разбивается и частично переходит с одного тела на другое. В теле создается избыток вихрей одного направления вращения. Пусть это будет вихри с вращением по часовой стрелке. Тело приобретет положительный заряд. Неспаренные «положительные» вихри будут вытолкнуты спаренными вихрями на поверхность тела, а уже на поверхности в соответствии с принципом наименьшего действия эти вихри равномерно будут распределены, естественно, с учетом радиуса кривизны поверхности тела, вихри распределяются только на выпуклых участках тела и плотность вихрей выше там, где радиус кривизны поверхности тела меньше.
Выдавленный (или созданный) на поверхность тела непарный вихрь при вращении будет за счет трения вовлекать во вращение прилегающий к телу жидкий эфир, заставляя его формировать вихрь. Возникший в жидком эфире вихревой поток будет напоминать гриб с большой шляпкой на тонкой ножке. Радиус «шляпки» такого вихря будет пропорционален квадрату расстояния от «центра» заряженного тела, поэтому «напряженность» в таком вихре будет падать обратно пропорционально квадрату расстояния (радиусу) от центра заряженного тела. Если теперь в пространстве, а точнее во все том же жидком эфире встретятся два таких вихря, то они начнут взаимодействовать. Вихри от «зарядов» с разными знаками будут «вкручиваться» друг в друга, а вихри от одноименных зарядов «выкручиваться друг из друга. Там, где вихревые потоки будут вкручиваться друг в друга, давление эфира будет снижаться, а там, где потоки будут «выкручиваться», давление эфира будет повышаться. В результате такого взаимодействия окружающий заряды жидкий эфир будет в первом случае заряды прижимать друг к другу, а во втором случае «эфирная подушка» с повышенным давлением в ней будет заряды раздвигать. И сила взаимодействия между зарядами будет прямопропорциональна произведению величин зарядов и обратно пропорциональна квадрату расстояния между зарядами.
Внимательный читатель обратил внимание, что нигде не идет речи о электронах, как своеобразной единице электричества. В этом пока нет никакой необходимости. Но тот факт, что электрические заряды состоят из неких единиц заставляет предположить, что в этом есть глубокий смысл, так уж устроен мир эфира, что единицей вихревого движения выступает некий стандартный параметр, как отражение неизвестных пока нам закономерностей. И с позиций эфирной теории электрона как такового, возможно, и нет, а есть некий эфирный вихрь, окружающий суммарный вихрь атома, и который может принимать в зависимости от обстоятельств самые разные формы как варианты объемной фигуры Лиссажу, некая стоячая эфирная волна из эфирной пены (эфирного пара), как реакция на колебания нуклонов или внешнее воздействие на сам атом. И этот окружающий атом или молекулу эфирный вихрь во многих случаях является носителем так называемой тепловой энергии. И чем больше такая «шуба» у атома или молекулы, тем больше тепловой энергии можно с такого атома или молекулы снять, если найти способ объединить такой атом или молекулу с другой и в результате новое соединение заставит исходные атомы или молекулы сбросить эфирную шубу, как мешающую этому объединению. Для создания такой шубы надо энергию где-то взять, а при исчезновении (разрушении) эфирной шубы, как варианта конденсации пара эфира в жидкий эфир энергия будет выделяться.
Например, атом водорода или кислорода окружены эфирной шубой. Но когда два атома водорода создают молекулу водорода, то они разворачиваются друг к другу так, чтобы направление вращения их вихрей было противоположным, т.е., создается электрически нейтральная спарка вихрей. Поэтому лишняя эфирная шуба будет сброшена за ненадобностью. А появление этой шубы в окружающем пространстве будет воспринято как повышение температуры. Также и с кислородом, когда он вступает в реакцию окисления, например, с углеродом. В результате создается электрически нейтральная молекула, которой компенсаторная эфирная шуба уже не нужна, сброшенный с кислорода и углерода эфир и идет на нагревание и освещение окружающего пространства. Так горит газ, бензин и все виды углеводородов, в которых кроме углерода очень много водорода. И когда 2 атома водорода встречаются с атомом кислорода, то все они сбрасывают с себя лишнюю защитную компенсирующую эфирную оболочку с выделением огромной энергии в виде светового и теплового излучения. Слава Богу, что так, что не возникает при этом гамма-излучения или атомных превращений с выбросом протонов и нейтронов. Именно на этих закономерностях можно создать источник энергии, работающий, например, исключительно на воде, которая будет использоваться в замкнутом цикле. Важно только правильно рассчитать, где атомы смогут обрести эфирную шубу за счет эфира окружающего пространства, а на каком этапе эту эфирную шубу можно с пользой для дела снять. Вот оно «Золотое руно». Надо еще проверить за каким собственно Золотым руном ездили аргонавты под руководством Ясона. Может быть, их послали за каким-нибудь альтернативным энергогенератором? Есть над чем подумать.
Теперь посмотрим, что такое магнитное поле. Так называемое магнитное поле – это тоже вихрь в жидком эфире, но особенностью такого вихря является то, что он уже создается не единичным зарядом, а за счет воздействия на жидкий эфир электростатического поля движущихся зарядов. Наиболее часто магнитное поле мы наблюдаем в постоянных магнитах. Это поле имеет часть, расположенную внутри магнита и часть – вне. Внутри магнита магнитный вихрь создается согласованным вращением атомов магнита или электронных оболочек атомов магнита. Ампер называл такие вращения элементарными токами. За счет синхронного вращения части атомов магнита, которые согласованно образуют эфирный насос, в объеме магнита формируются потоки жидкого эфира, вначале из того жидкого эфира, который есть в магните, как веществе, а затем уже магнит засасывает с одного полюса (южного) и выбрасывает с другого (северного) жидкий эфир окружающего пространства. И с того момента, как сформируются замкнутые магнитные силовые линии, похожие на винт Архимеда, можно считать, что магнитное поле магниту не принадлежит. Но это только в том случае, если мы будем поворачивать магнит вокруг оси симметрии. Но как только мы повернем магнит вокруг другой оси, как положение магнитного поля магнита соответствующим образом изменится. Магнитные силовые линии в виде «винтов Архимеда», но построенных из жидкого эфира, замкнуты сами на себя. И создается такое поле согласованным вращением токов Ампера, которые прогоняя жидкий эфир через себя, при этом эфир закручивают по часовой стрелке. Так что, фактически заряды, вращающие жидкий эфир существуют, но они расположены внутри магнита. И, наверное, поэтому Говард Джонс назвал такие токи неспаренными электронами, правильно определив характер вращения магнитного поля внутри магнита как результат вращения замкнутых токов или внутренних неспаренных зарядов. Можно даже предположить, что внутренние неспаренные заряды, фирменная особенность магнита. Так как, возникнув, такие заряды не могут по каким-то причина мигрировать на поверхность магнита, и не могут просто так исчезнуть. Надо еще вспомнить, как создаются постоянные магниты. А создаются они под воздействием на магнитную заготовку мощного электромагнитного импульса, который, возможно, разделяет атомные спарки (сдвоенные атомные вихри, вращающиеся в разные стороны) и часть их выбрасывает за пределы магнита. Оставшиеся неспаренные атомные вихри застревают и фиксируются внутри магнита в положении, которое задается направлением электромагнитного импульса. И сразу же принимаются перекачивать эфир с одного конца магнита к другому. И будут это делать, пока иная мощная причина не перемагнитит магнит или полностью лишит его магнитных свойств. И видимо в последнем случае неспаренные вихри внутри магнита будут дополнены вихрями с противоположным направлением вращения и так называемые токи Ампера прекратят свое существование.
Между движением жидкого эфира внутри постоянного магнита и движением постоянного тока в проводнике просматривается очень тесная связью. Только в первом случае циркуляция жидкого эфира создается неспаренными вихрями, созданными ранее в помощью электромагнитного импульса, а движением жидкого эфира в качестве постоянного тока обеспечивается динамически возникающими и пропадающими неспаренными вихрями при действии на проводник разности эфирного давления, которое в электротехнике называют напряжением. Чем больше напряжение, тем больше разрушается спаренных вихрей и образуется неспаренных вихрей, прогоняющих эфир от плюса к минусу. Механизм разрушения спаренных вихрей связан с удалением с части атомов некой эфирной оболочки, некого эфирного вихря, который слабо связан с ядрами атомов. Удаляемый и перемещающийся от атома к атому вихрь и носит название электрон и с которым связывают наличие электрического тока. Так и пишут, электрический ток – это упорядоченное движение электронов. А на деле движение электронов – это следствие прохождение по проводнику мощного потока жидкого эфира, поддерживаемого напряжением на концах проводника. И если проводник медный, то после снятия тока движение жидкого эфира по такому проводнику прекращается, а разрушенные вихри опять образуют электрически нейтральные пары вихрей. Но если проводник железный или стальной, короче ферромагнетик, то у такого проводника после снятия напряжения остается остаточная намагниченность. Т.е. пропуская по железной проволоке электрический ток, можно получить постоянный магнит. Этот факт отмечал в своих труде «Магнитный поток» Эд Лидскалнин, архитектор и единственный строитель Кораллового Замка, в котором общий вес блоков из кораллового известняка составляет 1100 тонн, а самые большие блоки доходят по весу под 50 тонн, и с такими блоками Эд Лидскалнин справлялся один без посторонней помощи и даже умудрялся просверливать в них идеальные отверстия длиной более трех метров.
Когда электрический ток пропускается по проводнику, то вращение потоков жидкого эфира в нём приводит к тому, что прилегаемый к проводнику жидкий эфир вовлекается во вращение и таким образом создается так называемое магнитное поле проводника с током. Это поле цилиндрическое, вращается в том же направление, что и поток эфира в проводнике, поэтому помещённый в магнитное поле проводник с током взаимодействует с этим током через взаимодействие магнитных полей. Магнитное поле проводника с током – это своеобразная спираль, намотанная на цилиндр, точнее много спиралей с разными радиусами, и если такая магнитная спираль помещается в однородное магнитное поле, то магнитное поле проводника с током вместе с самим проводником выталкивается из поля или перемещается в нём под действием силы, происхождение которой связано с эффектом Магнуса. Таким образом и на примере проводника с током мы убеждаемся в важности эффекта Магнуса.
Но еще Эд Лискалнин заметил, что между электрическим током и магнитным потоком внутри постоянного магнита нет никакой разницы. Материальный агент, который перемещается в проводнике с током и вдоль (внутри) постоянного магнита – это одна и та же материальная субстанция – жидкий эфир, закрученный в спираль с направлением вращения по часовой стрелке. Так что с эфирных позиций легко можно сказать, что такое электрический ток, и что между магнитным потоком и электрическим током разница заключается только в механизме создания эфирного потока.
Если постоянный магнит имеет осевую симметрию, то магнитный поток, выйдя из северного полюса, где давление эфира будет повышено, начнет двигаться к южному полюсу, где давление эфира будет понижено. В целом внутри и вне магнита возникнет торовидное осесемметричное магнитное поле. И даже, если мы поместим такой магнит в однородное магнитное поле другого более мощного магнита, то взаимодействовать такие магниты, а точнее их магнитные поля так, чтобы создавались силы, способные переместить магнит в магнитном поле более сильного магнита, не будут. Ибо сколько эфира будет проходить в единицу времени внутри магнита, столько же эфира будет двигаться в обратном направлении, но уже вокруг магнита. И вращаться вокруг оси такой магнит не будет по той же причине, ибо момент вращения эфира внутри магнита равен моменту вращения эфира вне магнита, но с противоположным знаком.
Чтобы создать условия для взаимодействия магнитных потоков (магнитных полей) надо понимать, что для потоков жидкого эфира справедливы те же законы, которым подчиняются потоки той же воды, только параметры у жидкого эфира другие, как практически несжимаемой жидкости, способной при некоторых режимах к парообразованию, т.е., кавитации. Для эфира кавитационный пузырёк, да еще и завихренный до скоростей на уровне скорости света и выше и есть вещество. И поэтому пока параметры электростатического или магнитного поля таковы, что эфир не превращается в пар, нет опасности превращения жидкого эфира в вещество. И, наоборот, при некоторых параметрах вихрей в жидком эфире создаются условия, при которых завихренный эфирный пар будет конденсироваться и это приведет к внезапному превращению вещества в жидкий эфир, что для наблюдателя будет похоже на испарение вещества в никуда. Но в данной статье нам нет необходимости рассматривать эти два крайних случая. Будем считать, что магнитное поле не нарушает структуру магнита, а магнит не приводит в изменению фазового состояния протекающего через него жидкого эфира.
Для того, чтобы получить возможность управлять магнитным потоком постоянного магнита, надо придать магниту такую форму, при которой магнитный поток преимущественно замыкался бы в кольцо. А для этого прямой магнит надо изогнуть, например, как это сделал Говард Джонс, используя для ротора (арматуры) своего мотора магниты в форме бумеранга.
Рис.1. Схема, поясняющая работу мотора Говарда Джонса.
Что произошло
Автор Дмитрий Иванович Менделеев
Оригинальная версия: Дата создания: октябрь 1902 г.,
...
Впервые опубликовано
«Попытка химического понимания мирового эфира», СПб., 1905, типолитография М. П. Фроловой, стр. 5—40.
Версия в PDF
Д. И. Менделеев «Периодический закон».
В редакции Б.М.Кедрова. Издательство Академии Наук СССР, Москва, 1958. с. 470 - 517.
Как рыба об лёд испокон веков билась мысль мудрецов в своем стремлении к единству во всём, то есть в искании «начала всех начал», но добилась лишь того, что все же должна признавать нераздельную, однако и не сливаемую, познавательную троицу вечных и самобытных: вещества (материи), силы (энергии) и духа, хотя разграничить их до конца, без явного мистицизма, невозможно. Различение и даже противоположение, еще нередко встречающееся в виде остатка от средних веков, лишь материального от духовного или — что того менее обще — лишь покоя от движения, не выдержало пытливости мышления, потому что выражает крайность и, главное, потому, что покоя ни в чем, даже в смерти, найти не удаётся, а духовное мыслимо лишь в абстракте, в действительности же познается лишь чрез материально ощущаемое, то есть в сочетании с веществом и энергиею, которая сама по себе тоже не сознаваема без материи, так как движение требует и предполагает движущееся, которое само по себе лишь мысленно возможно без всякого движения и называется веществом. Ни совершенно слить, ни совершенно отделить, ни представить какие-либо переходные формы для духа, силы и вещества не удается никому, кроме явных мистиков и тех крайних, которые не хотят ничего знать ни про что духовное: разум, волю, желания, любовь и самосознание. Оставим этим мистикам их дуализм, а обратим внимание на то, что вечность, неизменную сущность, отсутствие нового происхождения или исчезновения и постоянство эволюционных проявлений или изменений признали люди не только для духа, но и для энергии или силы, равно как и для материи или вещества. Научное понимание окружающего, а потому и возможность обладания им для пользы людской, а не для одного простого ощущения (созерцания) и более или менее романтического (то есть латинско-средневекового) описания, начинается только с признания исходной вечности изучаемого, как видно лучше всего над химиею, которая как чистая, точная и прикладная наука — ведет свое начало от Лавуазье, признавшего и показавшего «вечность вещества», рядом с его постоянной, эволюционною изменчивостью. Такое, еще во многом смутное, но все же подлежащее уже анализу понимание исходной троицы познания (вещество, сила и дух) составляет основу современного реализма, глубоко отличающегося как от древнего, так и от еще недавнего, даже еще до ныне распространенного унитарного материализма, который все стремится познать из вещества и его движения,[1] и от еще более древнего и также кое-где еще не забытого унитарного же спиритуализма, все как будто понимающего, исходя из одного духовного. Думаю даже, что современный «реализм» яснее и полнее всего характеризуется признанием вечности, эволюции и связей: вещества, сил и духа.
Так, сколько я понимаю, мыслят вдумчивые естествоиспытатели — реалисты,[2] и это их в некоторой мере успокаивает, когда они изучают вещество, его формы и силы, в нем действующие, и когда они стремятся узнать их предвечные закономерности. Но у них есть свои побочные причины постоянного беспокойства. Их много. Одну из них выбираю предметом статьи, а именно мировой эфир, или просто «эфир». В известной краткой энциклопедии Ларусса (Pierre Larousse, Dictionnaire complet illustré), составляющей в некотором смысле экстракт и перечень современно-известного и признанного, вот как определяется «эфир» (éther): «жидкость невесомая, упругая, наполняющая пространство, проникающая во все тела и признаваемая физиками за причину света, тепла, электричества и проч.».
Сказано немного, но достаточно для того, чтобы смущать вдумчивых естествоиспытателей. Они не могут не признать за эфиром свойств вещества (здесь «жидкости»), а в то же время придумали его, как мировую «среду», наполняющую всё пустое пространство и все тела, чтобы уразуметь хоть сколько-нибудь при помощи движения этой среды передачу энергии на расстояния, и признали в этой среде разнообразные перемены строения (деформации) и возмущения (пертурбации), какие наблюдаются в твердых телах, жидкостях и газообразных веществах, чтобы ими толковать явления света, электричества и даже тяготения. В этой жидкой среде нельзя показать весомости, если эта жидкость всюду и всё проникает, как нельзя было знать весомости воздуха, пока не нашли воздушных насосов, способных удалять воздух. Но нельзя и отрицать весомости эфира, потому что со времён Галилея и Ньютона способность притягиваться, то есть весить, составляет первичное определение вещества. Путем совокупности предположений В. Томсон (лорд Кельвин) пришёл к выводу, что кубический метр эфира должен весить, примерно, не менее 0,000 000 000 000 000 1 грамма, если куб. метр воды весит около 1 000 000 граммов,[3] а для легчайшего — водородного — газа при 0° и при обыкновенном атмосферном давлении куб. метр весит около 90 граммов. В совершенно законном стремлении придать эфиру весомость или массу начинается то беспокойство вдумчивых естествоиспытателей, о котором сказано выше, потому что рождается вопрос: да при каком же давлении и при какой же температуре эфиру свойствен указанный вес? Ведь, и для воды и водорода при ничтожно малых давлениях или при громадных повышениях температуры должно ждать такой же малой плотности, какая выше указана для эфира. Если дело идет о плотности эфира в междупланетном пространстве, то там и водяные пары, и водород не могут иметь, несмотря на низкую температуру, видимой, измеримой плотности, так как там давления, определяемые тяготением, ничтожны. Умственно можно представить, что междупланетное пространство наполнено такими разреженными остатками всяких паров и газов. Даже тогда получится согласие с известными космогоническими гипотезами Канта, Лапласа и др., стремящимися выяснить единство плана образования миров, поймется однообразие химического состава всей вселенной, указанное спектрометрическими исследованиями, так как по существу установится обмен — чрез посредство эфира — между всеми мирами. Исследование упругости или сжимаемости газов под малыми давлениями, задуманное мною в 70-х годах и отчасти тогда же выполненное, имело, между прочим, целью проследить, насколько то возможно для имеющихся способов измерений малых давлений, изменения в газах, находящихся под малыми давлениями. Подмеченные для всех газов (мною с М. Л. Кирпичевым, 1874) так называемые положительные отступления от Бойль—Мариоттова закона, затем подтвержденные многими и, между прочим, Рамзаем (хотя до сих пор и непризнаваемые еще некоторыми исследователями), до некоторой степени указывают на единообразие поведения всех газов и на стремление их при уменьшении давления к некоторому пределу в расширении, как есть предел для сгущения — в сжижении и критическом состоянии.[4] Но в наблюдении очень малых давлений встретились непреоборимые трудности, тем большие, что для определения очень малых давлений оказалось невозможным заменить ртуть более легкими жидкостями (например, серною кислотою или нефтяными маслами), потому что они оказались способными выделять из себя в манометрическую пустоту ничтожно малые, однако ясно видимые количества каких-то газов, хотя были предварительно неделями выдержаны при 100° в пустоте, доставляемой лучшими насосами. Таким образом практически оказалось невозможным сколько-либо точно измерять давления, меньшие, чем в десятые доли миллиметра высоты ртутного столба, а это — когда дело идет о разрежениях, подобных тем, какие надо предполагать даже на высоте 50 километров над уровнем наших морей — чересчур большие величины. Поэтому представление об эфире как сильно-разреженном газе атмосферы, не может доныне подлежать опытному исследованию и измерению, которые одни способны наводить (индуцировать) мысль на правильные пути и приводить затем к следствиям, опять подлежащим опытной и измерительной поверке.
Но и помимо этого, представление о мировом эфире, как предельном разрежении паров и газов, не выдерживает даже первых приступов вдумчивости — в силу того, что эфир нельзя представить иначе, как веществом, все и всюду проникающим; парам же и газам это не свойственно. Они сгущаемы при увеличении давлений, и их нельзя представить содержащимися во всех веществах, хотя они и широко распространены во всех телах природы, даже в аэролитах. Притом — и это, всего важнее — они, по своей химической природе и по своим отношениям к другим веществам, бесконечно разнообразны; эфир же однообразен всюду, насколько то нам известно. Будучи разнородны по своим химическим свойствам, известные нам пары и газы должны были бы химически разнообразно воздействовать на тела, которые они проникают, если бы эфир был их совокупностью.
Прежде чем итти далее, считаю неизбежно необходимым оговориться в отношении здесь и далее вводимых мною химических соображений. Избежать их при обсуждении мирового эфира было трудно, но во времена Галилея и Ньютона еще возможно. Ныне же это было бы противно самым основным началам дисциплины естественной философии, потому что со времен Лавуазье, Дальтона и Авогадро—Жерара химия получила все высшие права гражданства в обществе наук о природе и, поставив массу (вес) вещества во главе всех своих обобщений, пошла за Галилеем и Ньютоном. Мало того, чрез химию, только при ее приемах, действительно вкоренилось во всем естествознании стремление искать решение всяких задач, касающихся конечных, измеримых тел и явлений, в постижении взаимодействия беспредельно малых их отдельностей, называемых атомами, но в сущности (по реальному представлению) мыслимых, как химически неделимые индивидуумы, ничего общего не имеющих с механически-неделимыми атомами древних метафизиков. Доказательства этому последнему многочисленны, но достаточно упомянуть о том, что современные атомы не раз объясняли вихревыми кольцами (vortex), что и поныне живо стремление понять сложение химических атомов или друг из друга, или из «первичной материи» и что как раз в последнее время, особенно по поводу радиоактивных веществ, стали признавать деление химических атомов на более мелкие «электроны», а все это логически не было бы возможно, если бы «атомы» признавались механически неделимыми. Химическое миросозерцание можно выразить образно, уподобляя атомы химиков небесным телам: звездам, солнцу, планетам, спутникам, кометам и т. п. Как из этих отдельностей (индивидуумов) слагаются системы, подобные солнечной или системам двойных звёзд, или некоторым созвездиям (туманностям) и т. п., так представляется сложение из атомов целых частиц, а из частиц тел и веществ. Это для современной химии не простая игра слов или не одно уподобление, а сама реальность, руководящая всеми исследованиями, всякими анализами и синтезами химии. У нее свой микрокосм в невидимых областях, и, будучи архиреальною наукою, она все время оперирует с невидимыми своими отдельностями, вовсе не думая считать их механически неделимыми. Атомы и частицы (молекулы), о которых неизбежно говорится во всех частях современной механики и физики, не могут быть чем-либо иным, как атомами и частицами, определяемыми химией, потому что того требует единство познания. Поэтому и метафизика нашего времени, если желает помогать познанию, должна понимать атомы так же, как их понимать могут естествоиспытатели, а не на манер древних метафизиков китайско-греческого образца. Если Ньютоново всемирное тяготение реально раскрыло силы, всегда действующие даже на беспредельно больших расстояниях, то познание химии, внушенное Лавуазье, Дальтоном и Авогадро—Жераром, раскрыло силы, всегда действующие на неизмеримо малых расстояниях, и показало как громадность этих сил (что видно, например, из того, что силами этими легко сжижаются газы, подобные водороду, едва недавно сжиженному совокупностью физических и механических усилий), так и превращаемость их во все прочие виды проявления энергии, так как химическими силами (например при горении) достигаются механические и физические. Поэтому все современные основные понятия естествознания — следовательно, и мировой эфир — неизбежно необходимо обсудить под совокупным воздействием сведений механики, физики и химии, и, хотя понятие об эфире родилось в физике, и хотя скептическая индифферентность старается во всем усмотреть «рабочую гипотезу», вдумчивому естествоиспытателю, ищущему саму действительность, какова она есть, и не довольствующемуся смутными картинами волшебного фонаря фантазии, хотя бы украшенного логичнейшим анализом, нельзя не задаваться вопросом: что же такое это за вещество в химическом смысле?
Моя попытка и начинается с этого вопроса.
Ранее, чем излагать свой посильный ответ на вопрос о химической природе эфира, считаю долгом высказаться о мнении, которое читал между строк и не раз слыхал от своих учёных друзей, верящих в единство вещества химических элементов (или простых тел) и в происхождение их из одной первичной материи. Для них эфир содержит эту первичную материю в несложившемся виде, то есть не в форме элементарных химических атомов и образуемых ими частиц и веществ, а в виде составного начала, из которого сложились сами химические атомы. Нельзя не признать в таком воззрении увлекательной стороны. Как миры представляют иногда сложившимися из разъединенных тел (твердой космической пыли, болидов и т. п.), так атомы представляют происшедшими из первичного вещества. Сложившиеся миры остаются, но рядом с ними остается в пространстве космическая пыль, кометы, болиды и т. п. материалы, из которых предполагается их сложение уже многими. Так остаются и сложившиеся атомы, но рядом с ними сохранился и между ними движется их материал, то есть всепроникающий и первозданный эфир. Одни при этом полагают, что есть ряд видимых явлений, при которых атомы рассыпаются в свою пыль, то есть в первичную материю, как рассыпаются кометы в потоки падающих звезд. Химики и физики, так думающие, представляют, что как геологические изменения или как сложение и распадение миров идут перед нашими глазами, так пред нами же в тиши разрушаются и вновь слагаются и атомы в своей вечной эволюции. Другие, не отрицая такой возможности — в виде особо редкого и исключительного случая, считают мир атомов сложенным в твердь прочно и полагают невозможным направить опыт на то, чтобы уловить это, то есть считают невозможным на опыте рассыпать атомы в первичную материю или образовать из неё на наших глазах новые атомы химических элементов, то есть процесс их происхождения понимают раз бывшим и законченным навсегда, а в эфире видят остатки, отбросы. С последними — реалистами не приходится считаться, потому что при таком представлении мыслители руководятся не следствиями из наблюдений или опытов, а только воображением, свобода которого обеспечена в республике науки. Но с первыми, то есть с истинными поклонниками продолжающейся эволюции вещества атомов, считаться химическому реализму неизбежно, потому что исходные положения нашей науки состоят не только в том, что вся общая масса вещества постоянна, но постоянны и те формы вещества, которые понимаются как элементарные атомы и в отдельности являются как «тела простые», признаваемые неспособными превращаться друг в друга. Если бы эфир происходил из атомов и атомы из него слагались, то нельзя было бы отрицать образование новых, небывалых атомов и должно было бы признавать возможность исчезания части простых тел, взятых в дело, при тех или иных наблюдениях и опытах. Давно- давно масса людей, по старому предрассудку, верит в такую возможность и, если бы это мнение не сохранялось в наши дни, не являлись бы Емменсы в С. А. С. Штатах, стремящиеся, по манере алхимиков, превратить серебро в золото, или такие учёные, как Фиттика (F. Fittica), в Германии, который еще недавно, в 1900 году, старался доказывать, что фосфор может превращаться в мышьяк. Множество случаев подобного превращения одних простых тел в другие описывалось в те 50 лет, в течение которых я внимательно слежу за химической литературой. Но каждый раз, при тщательном исследовании подобных случаев, оказывалась или простая ошибка предубеждения, или недостаточная точность исследования, и вновь[5] защищать индивидуальную самобытность химических элементов я здесь не предполагаю. Мне следовало, однако, напомнить об этом, рассматривая эфир, потому что, помимо химической бездоказательности, мне кажется, невозможно сколько-либо реальное понимание эфира, как первичного вещества, потому что у веществ первейшими принадлежностями должно считать массу или вес и химические отношения: — первую для понимания большинства явлений при всех расстояниях, вплоть до бесконечно больших, а вторые — при расстоянии неизмеримо малых или соизмеримых с величинами тех мельчайших отдельностей, которые называют атомами. Если бы дело шло об одном том эфире, который наполняет пространство между мировыми телами (солнцем, планетами и т. п). и передает между ними энергию, то можно было бы — с грехом пополам, ограничиваться только предположением о массе, не касаясь его химизма, можно было бы даже считать эфир содержащим «первичную материю», как можно говорить о массе планеты, не касаясь ее химических составных начал. Но вполне, так сказать, бескровный, ближе ничем не определяемый эфир окончательно теряет всякую реальность и составляет причину беспокойства вдумчивых естествоиспытателей, лишь только спускаемся с неба на землю и признаем его проникающим все тела природы. Необходимость лёгкого и полного проникновения всех тел эфиром следует признать не только ради возможности понимания множества общеизвестных физических явлений, начиная с оптических (над чем не считаю надобным останавливаться), но и по причине великой упругости и, так сказать, тонкости эфирного вещества, атомы которого всегда и все представляют себе не иначе, как очень малыми сравнительно с атомами и частицами химически известных веществ, то есть подобными аэролитам среди планет. Притом такая проницаемость эфиром всех тел объясняет и невозможность уединить это вещество, как нельзя собрать ни воды, ни воздуха в решете, каким для эфира должно считать всякие твердые или иные вещества и преграды. Способность эфира проникать всюду, во все тела можно, однако, понимать, как высшую степень развития того проникновения газов чрез сплошные преграды, которое Грем изучал для каучука в отношении многих газов, а Девилль и др. нашли для железа и платины по отношению к водороду[6].
Обладая малым весом атома и низшею из всех известных газов плотностью, водород не только вытекает или диффундирует сильнее или быстрее всяких других газов чрез малейшие отверстия, но способен проникать и чрез сплошные стенки таких металлов, как платина и особенно палладий, чрез которые другие газы не проникают. Но тут несомненно действует не только быстрота движения частиц водорода, тесно связанная с его малою плотностью, но и химическая способность того же разряда, которая проявляется как при образовании сложных тел, содержащих водород, так и при образовании растворов, сплавов и тому подобных, так называемых, неопределенных соединений. Механизм этого проникновения можно представить подобным — на поверхности проницаемого тела — растворению газа в жидкости, то есть вскакиванию его частиц в промежутки между частицами жидкости, замедлению движения (отчасти некоторому сгущению газа) и такому или иному согласованию движений обоих видов частиц. В массе проницаемого тела сжатый газ, поглощенный на поверхности прикосновения, конечно, распространяется во все стороны, диффундируя от слоя к слою, если в опытах Робертс-Аустена даже золото диффундировало в твердом свинце на основании тех же сил. Наконец, на другой поверхности проницаемого тела сжатый газ находит возможность вырваться на большую свободу и, пока будет накопляться до исходного давления, станет проникать туда, где его нет или где его мало, то есть входить в преграду будет более со стороны превышающего давления, чем в обратном направлении. Когда же давления уравняются, наступит не покой, а подвижное равновесие, то есть с каждой стороны в преграду будет проникать и выбывать одинаковое число частиц или атомов. Допуская, а это необходимо, проницаемость эфира в отношении ко всем веществам, должно приписать ему, прежде всего, лёгкость и упругость, то есть быстроту собственного движения, еще в большем развитии, чем для водорода, и, что всегда важнее, ему должно приписать еще меньшую, чем для водорода, способность образовать с проницаемыми телами определённые химические соединения, так как эти последние характеризуются именно тем, что разнородные атомы образуют системы или частицы, в которых вместе или согласно движутся различные элементы, как солнечная система характеризуется зависимым, согласным и совместным движением образующих ее многих светил. А так как надо предполагать, что такое совместное движение водорода, например, с палладием, им проницаемым, действительно совершается для тех атомов водорода, которые находятся в среде атомов палладия, и что водород с палладием дает своё определённое соединение Pd²H (или какое иное), но при нагревании оно легко диссоциирует, то следует, мне кажется, допустить, что атомы эфира в такой высокой мере лишены этой, уже для водорода слабой, способности к образованию определённых соединений, что для них всякая температура есть диссоциационная, а потому ничего, кроме некоторого сгущения в среде атомов обычного вещества, для эфира признать нельзя.
Такое допущение, то есть отрицание для вещества или для атомов эфира всякой склонности к образованию сколько-либо стойких соединений с другими химическими элементами, ещё несколько лет тому назад должно было бы считать совершенно произвольным, а потому и мало вероятным даже гипотетически, так как все известные еще недавно простые тела и элементы, так или иначе, труднее или легче и прочнее или шатче, прямо или косвенно вступали во взаимные соединения, и тогда представить вещество, вовсе лишенное склонности подвергнуться под влиянием других веществ каким-либо химическим изменениям и чуждое способности образовать сложные частицы, — было бы чересчур смело и лишено всякой реальности, то есть чуждо известной действительности. Но вот в 1894 г. лорд Релей и проф. Рамзай открывают в воздухе аргон и определяют его, как недеятельнейшее из всех известных газообразных и всяких иных веществ. Скоро затем последовало открытие Рамзаем гелия, который по его яркому спектру Локьер предчувствовал, как особое простое тело на солнце; а затем Рамзай и Траверс открыли в сжиженном воздухе еще три таких же недеятельных, как аргон, газа: неон, криптон и ксенон, хотя содержание их в воздухе ничтожно мало и должно считаться для гелия и ксенона миллионными долями по объему и весу воздуха[7]. Для этих пяти новых газов, составляющих, вместе с открытием радиоактивных веществ, одни из блистательнейших опытных открытий конца XIX века, до сих пор не получено никаких сложных соединений, хотя в них ясно развита способность сжижаться и растворяться, то есть образовать так называемые неопределённые, столь легко диссоциирующие, соединения. Поэтому ныне, с реальной точки зрения, уже смело можно признавать вещество эфира лишённым — при способности проникать все вещества — способности образовать с обычными химическими атомами какие-либо стойкие химические соединения. Следовательно, мировой эфир можно представить, подобно гелию и аргону, газом, не способным к химическим соединениям.
Оставаясь на чисто химической почве, мы старались сперва показать невозможность понимания эфира ни как рассеянный пар или газ всюду распространенных веществ, ни как атомную пыль первичного вещества, из которого нередко еще доныне многие при знают сложение элементарных атомов, а потом пришли к заключению о том, что в эфире должно видеть вещество, лишённое способности вступать в сколько-либо прочные определенные химические соединения, что свойственно недавно открытым гелию, аргону и их аналогам.
Это первый этап на нашем пути; на нем, хотя недолго, необходимо остановиться. Когда мы признаем эфир газом — это значит прежде всего, что мы стремимся отнести понятие о нём к обычным, реальным понятиям о трёх состояниях веществ: газообразном, жидком и твердом. Тут не надо признавать, как то делает Крукс, особого четвёртого состояния, ускользающего от реального понимания природы вещей. Таинственная, почти спиритическая подкладка с эфира при этом допущении скидывается. Говоря, что это есть газ, очевидно, мы признаём его «жидкостью» в широком смысле этого слова, так как газы вообще суть упругие жидкости, лишённые сцепления, то есть той способности настоящих жидкостей, которая проявляется в виде свойства образовать — в силу сцепления — капли, подниматься в волосных (капиллярных) трубках и т. п. У жидкостей мера сцепления есть определённая, конечная величина, у газов она близка к нулю или, если угодно, величина очень малая. Если эфир — газ, то, значит, он имеет свой вес; это неизбежно приписать ему, если не отвергать ради него всей концепции естествознания, ведущего начало от Галилея, Ньютона и Лавуазье. Но если эфир обладает столь сильно развитою проницаемостью, что проходит чрез всякие оболочки, то нельзя и думать о том, чтобы прямо из опыта найти его массу в данном количестве других тел, или вес его определённого объема — при данных условиях, а потому должно говорить не об невесомом эфире, а только о невозможности его взвешивания. Конечно, тут скрыта своя гипотеза, но совершенно реальная, а не какая-то мистическая, внушающая сильное беспокойство вдумчивым естествоиспытателям.
Всё предшествующее, мне кажется, не только не противоречит общераспространённому представлению о мировом эфире, но прямо с ним согласуется. Добавка, нами сделанная, стремящаяся ближе реализовать понятие об эфире, состоит только в том, что мы пришли к необходимости и возможности приписать эфиру свойства газов, подобных гелию и аргону, и в наивысшей мере неспособность вступать в настоящие химические соединения. Над этим понятием, составляющим центральную посылку моей попытки, необходимо остановиться подробнее, чем над какою-либо иною стороною сложного и важного предмета, например, над сопротивлением эфирной среды движению небесных светил, над следованием за Бойль—Мариоттовым или Ван-дер-Ваальсовым законом, над громадною упругостью массы эфира, над мерою его сгущения и упругостью в разных телах и в небесном пространстве и т. п. Все такие вопросы придётся так или иначе умственно решать и при всяком ином представлении об эфире, как весомом, но не взвешиваемом веществе. Мне кажутся все эти стороны доступными для реального обсуждения уже ныне, но они завлекли бы нас слишком далеко и все же основной вопрос — о химическом составе эфира — остался бы при этом висеть в пустоте, а без него не может быть, на мой взгляд, никакой реальности в суждении об эфире; после же такого или иного ответа на этот вопрос, быть может, удастся двинуться дальше в реальном понимании других отношений эфира. Поэтому далее я стану говорить только о своей попытке понять химизм эфира, исходя из двух основных положений, а именно: 1) эфир есть легчайший — в этом отношении предельный — газ, обладающий высокою степенью проницаемости, что в физико-химическом смысле значит, что его частицы имеют относительно малый вес и обладают высшею, чем для каких-либо иных газов, скоростью своего поступательного движения, [8] и 2) эфир есть простое тело, лишённое способности сжижаться и вступать в частичное химическое соединение и реагирование с какими-либо другими простыми или сложными веществами, хотя способное их проницать, подобно тому, как гелий, аргон и их аналоги способны растворяться в воде и других жидкостях.
Дальнейшие стороны моей попытки — понять природу эфира — так тесно связаны с гелием, аргоном и их аналогами и с периодическою системою элементов, что мне ранее, чем итти вперед, необходимо особо остановиться над этими предметами и их взаимною связью.
Когда, в 1869 г., на основании сближений, подмеченных уж Дюма, Ленсеном, Петтенкофером и другими, между величинами атомных весов сходственных элементов, мною была выставлена периодическая зависимость между свойствами всех элементов и их истинными (то есть по системе Авогадро—Жерара с дополнениями Канницаро и с изменениями, вызываемыми периодическою законностью) атомными весами, не только не было известно ни одного элемента, неспособного образовать определенные сложные соединения, но нельзя было даже и подозревать возможности существования подобных элементов. Поэтому в периодической системе, данной мною в том виде, какой она сохранила и до сих пор, а именно при расположении по группам, рядам и периодам (см. 1-е издание книги моей «Основы химии», выпуск 3-й, вышедший в 1870 году, и статьи мои в журнале Русского Химического Общества 1869 г.), система элементов начиналась с группы 1-й и с ряда 1-го, где помещался и до сих пор помещается водород, легчайший из элементов, судя по атомному весу, и легчайший газ, судя по плотности, — при данных давлении и температуре. Никогда мне в голову не приходило, что именно водородом должен начинаться ряд элементов, хотя легче его не было и еще поныне между известными нет ни одного другого элементарного или сложного газа. Оставаясь на реальной почве, я решался предсказывать не только существование неизвестных элементов в среде известных, но и их свойства, как химические, так и физические, для них самих в свободном состоянии (простых тел) и для их соединений. Это, как известно, оправдалось последующими открытиями: галлия — Лекоком де Боабодраном, скандия — Нильсоном и, блистательнее всего, германия — Клементом Винклером, моим (ныне уже скончавшимся) хорошим другом и научным собратом. Предсказания эти были, по существу, тем, что называется в математике интерполированием, то есть нахождением, промежуточных точек на основании крайних, когда известен закон (или направление кривой, его выражающей), по которому точки следуют друг за другом. Поэтому оправдание предсказанного есть не что иное, как способ утверждения законности, и, следовательно, теперь можно смело полагаться на то, что в 1869—1871 гг. было только вероятным, и уверенно признавать, что химические элементы и их соединения находятся в периодической зависимости от атомных весов элементов. Эксполировать, то есть находить точки вне пределов известного, нельзя было на основании еще неупроченной законности. Но когда она утвердилась, можно на это решиться, и то, что дальше будет сказано об эфире, как элементе, гораздо более лёгком, чем водород, составляет такое эксполирование. Решимость моя, при той осторожности, какая должна быть свойственна всякому деятелю науки, определяется двумя соображениями. Во-первых, я думаю, что откладывать — по старости лет — мне уже нельзя. А, во-вторых, за последнее время стали много и часто говорить о раздроблении атомов на более мелкие электроны, а мне кажется, что такое дробление должно считать не столько метафизическим, сколько метахимическим представлением, вытекающим из отсутствия каких-либо определенных соображений, касающихся химизма эфира, и мне захотелось на место каких-то смутных идей поставить более реальное представление о химической природе эфира, так как, пока что-нибудь не покажет либо превращения обычного вещества в эфир и обратно, либо превращения одного элемента в другой, всякое представление о дроблении атомов должно считать, по моему мнению, противоречащим современной научной дисциплине, а те явления, в которых признаётся дробление атомов, могут быть понимаемы, как выделение атомов эфира, всюду проникающего и признаваемого всеми. Словом, мне кажется, хотя рискованным, но своевременным говорить о химической природе эфира, тем более, что, сколько мне известно, об этом предмете еще никто не говорил более или менее определённо. Когда я прилагал периодический закон к аналогам бора, алюминия и кремния, я был на 33 года моложе, во мне жила полная уверенность, что рано или поздно предвидимое должно непременно оправдаться, потому что мне все там было ясно видно. Оправдание пришло скорее, чем я мог надеяться. Теперь же у меня нет ни прежней ясности, ни бывшей уверенности. Тогда я не рисковал, теперь рискую. На это надобна решимость. Она пришла, когда я видел радиоактивные явления, как объяснено в конце статьи, и когда я сознал, что откладывать мне уже невозможно и что, быть может, мои несовершенные мысли наведут кого-нибудь на путь более верный, чем тот возможный, какой представляется моему слабеющему зрению.
Первоначально я выскажусь о положении гелия, аргона и их аналогов в периодической системе элементов, потом о представляемом мною месте эфира в той же системе, а закончу несколькими беглыми замечаниями по поводу ожидаемых свойств эфира, основанных на понятии о нём, выводимом из его положения в этой системе.
Когда в 1895 г. дошли до меня первые сведения об аргоне и его беспримерной химической инертности (он ни с чем, ни при каких условиях не реагирует), мне казалось законным сомневаться в элементарной простоте этого газа, и я предполагал, что аргон можно считать полимером азота N³, как озон О³ есть полимер кислорода О² , но с тем различием, что озон происходит, как известно, из кислорода с присоединением — как говорится — тепла, то есть выделяет на данный свой вес более тепла, вступая в реакции, одинаковые с кислородом, чем кислород при том же весе, а аргон можно было представить, как азот, потерявший тепло, то есть еще менее энергичный, чем обычный азот. Этот последний всегда служил в химии образцом химической инертности, то есть простым телом, очень трудно вступающим в реакции, и если бы представить, что его атомы, уплотняясь при полимеризации из N² в N³ , теряют теплоту, можно было ждать вещества еще в высшей мере инертного, то есть ещё более сопротивляющегося воздействию других веществ. Так, кремнезём, происходящий с отделением тепла из кремния и кислорода, менее последних способен к химическим реакциям. Подобное же представление о природе аргона и о связи его с азотом высказано было затем известнейшим ученым Вертело. Теперь, уже давно, я отказался от такого мнения о природе аргона и соглашаюсь с тем, что это есть самостоятельное элементарное вещество, как это с самого начала утверждал Рамзай. Поводов к такой перемене было очень много. Главнейшими служили: 1) несомненная уверенность в том, что плотность аргона гораздо менее 21, а именно, вероятно, лишь немногим более 19, если плотность водорода принять за 1, а для N³ надо ждать плотности около 21, так как вес частицы N³ =3·14=42, а плотность близка к половине веса частицы; 2) гелий, открытый тем же Рамзаем в 1895 г., представляет плотность, по водороду, около 2-х и обладает такою же полною химическою инертностью, как и аргон, а для него нельзя уже было реально мыслить о сложности частицы и ею объяснять инертность; 3) такую же инертность Рамзай и Траверс нашли для открытых ими неона, криптона и ксенона, и что пригодно было для аргона — было не применимо к ним; 4) самостоятельные особенности спектра каждого из указанных пяти газов, при полной их неизменности от ряда электрических искр, убеждали, что это целая семья элементарных газов, глубоко отличающихся от всех, до тех пор известных, своею полною химическою инертностью, и 5) постепенность и определенность физических свойств в зависимости от плотности и от веса атома[9] дополняют, благодаря трудам того же Рамзая, уверенность в том, что здесь дело идет о простых телах, самобытность которых, при отсутствии химических превращений, и можно было утверждать только постоянством физических признаков. Укажем для примера на изменение температуры кипения (при давлении в 760 миллим.) или той, при которой достигается упругость, равная атмосферной, и могут существовать — при указанном давлении — как жидкая, так и газообразная фазы:
Химич. знак и состав частицы Гелий.
He Не
Попытка химического понимания мирового эфира
2012-04-29 17:45:00 (читать в оригинале)Автор Дмитрий Иванович Менделеев
Оригинальная версия: Дата создания: октябрь 1902 г.,
...
Впервые опубликовано
«Попытка химического понимания мирового эфира», СПб., 1905, типолитография М. П. Фроловой, стр. 5—40.
Версия в PDF
Д. И. Менделеев «Периодический закон».
В редакции Б.М.Кедрова. Издательство Академии Наук СССР, Москва, 1958. с. 470 - 517.
Как рыба об лёд испокон веков билась мысль мудрецов в своем стремлении к единству во всём, то есть в искании «начала всех начал», но добилась лишь того, что все же должна признавать нераздельную, однако и не сливаемую, познавательную троицу вечных и самобытных: вещества (материи), силы (энергии) и духа, хотя разграничить их до конца, без явного мистицизма, невозможно. Различение и даже противоположение, еще нередко встречающееся в виде остатка от средних веков, лишь материального от духовного или — что того менее обще — лишь покоя от движения, не выдержало пытливости мышления, потому что выражает крайность и, главное, потому, что покоя ни в чем, даже в смерти, найти не удаётся, а духовное мыслимо лишь в абстракте, в действительности же познается лишь чрез материально ощущаемое, то есть в сочетании с веществом и энергиею, которая сама по себе тоже не сознаваема без материи, так как движение требует и предполагает движущееся, которое само по себе лишь мысленно возможно без всякого движения и называется веществом. Ни совершенно слить, ни совершенно отделить, ни представить какие-либо переходные формы для духа, силы и вещества не удается никому, кроме явных мистиков и тех крайних, которые не хотят ничего знать ни про что духовное: разум, волю, желания, любовь и самосознание. Оставим этим мистикам их дуализм, а обратим внимание на то, что вечность, неизменную сущность, отсутствие нового происхождения или исчезновения и постоянство эволюционных проявлений или изменений признали люди не только для духа, но и для энергии или силы, равно как и для материи или вещества. Научное понимание окружающего, а потому и возможность обладания им для пользы людской, а не для одного простого ощущения (созерцания) и более или менее романтического (то есть латинско-средневекового) описания, начинается только с признания исходной вечности изучаемого, как видно лучше всего над химиею, которая как чистая, точная и прикладная наука — ведет свое начало от Лавуазье, признавшего и показавшего «вечность вещества», рядом с его постоянной, эволюционною изменчивостью. Такое, еще во многом смутное, но все же подлежащее уже анализу понимание исходной троицы познания (вещество, сила и дух) составляет основу современного реализма, глубоко отличающегося как от древнего, так и от еще недавнего, даже еще до ныне распространенного унитарного материализма, который все стремится познать из вещества и его движения,[1] и от еще более древнего и также кое-где еще не забытого унитарного же спиритуализма, все как будто понимающего, исходя из одного духовного. Думаю даже, что современный «реализм» яснее и полнее всего характеризуется признанием вечности, эволюции и связей: вещества, сил и духа.
Так, сколько я понимаю, мыслят вдумчивые естествоиспытатели — реалисты,[2] и это их в некоторой мере успокаивает, когда они изучают вещество, его формы и силы, в нем действующие, и когда они стремятся узнать их предвечные закономерности. Но у них есть свои побочные причины постоянного беспокойства. Их много. Одну из них выбираю предметом статьи, а именно мировой эфир, или просто «эфир». В известной краткой энциклопедии Ларусса (Pierre Larousse, Dictionnaire complet illustré), составляющей в некотором смысле экстракт и перечень современно-известного и признанного, вот как определяется «эфир» (éther): «жидкость невесомая, упругая, наполняющая пространство, проникающая во все тела и признаваемая физиками за причину света, тепла, электричества и проч.».
Сказано немного, но достаточно для того, чтобы смущать вдумчивых естествоиспытателей. Они не могут не признать за эфиром свойств вещества (здесь «жидкости»), а в то же время придумали его, как мировую «среду», наполняющую всё пустое пространство и все тела, чтобы уразуметь хоть сколько-нибудь при помощи движения этой среды передачу энергии на расстояния, и признали в этой среде разнообразные перемены строения (деформации) и возмущения (пертурбации), какие наблюдаются в твердых телах, жидкостях и газообразных веществах, чтобы ими толковать явления света, электричества и даже тяготения. В этой жидкой среде нельзя показать весомости, если эта жидкость всюду и всё проникает, как нельзя было знать весомости воздуха, пока не нашли воздушных насосов, способных удалять воздух. Но нельзя и отрицать весомости эфира, потому что со времён Галилея и Ньютона способность притягиваться, то есть весить, составляет первичное определение вещества. Путем совокупности предположений В. Томсон (лорд Кельвин) пришёл к выводу, что кубический метр эфира должен весить, примерно, не менее 0,000 000 000 000 000 1 грамма, если куб. метр воды весит около 1 000 000 граммов,[3] а для легчайшего — водородного — газа при 0° и при обыкновенном атмосферном давлении куб. метр весит около 90 граммов. В совершенно законном стремлении придать эфиру весомость или массу начинается то беспокойство вдумчивых естествоиспытателей, о котором сказано выше, потому что рождается вопрос: да при каком же давлении и при какой же температуре эфиру свойствен указанный вес? Ведь, и для воды и водорода при ничтожно малых давлениях или при громадных повышениях температуры должно ждать такой же малой плотности, какая выше указана для эфира. Если дело идет о плотности эфира в междупланетном пространстве, то там и водяные пары, и водород не могут иметь, несмотря на низкую температуру, видимой, измеримой плотности, так как там давления, определяемые тяготением, ничтожны. Умственно можно представить, что междупланетное пространство наполнено такими разреженными остатками всяких паров и газов. Даже тогда получится согласие с известными космогоническими гипотезами Канта, Лапласа и др., стремящимися выяснить единство плана образования миров, поймется однообразие химического состава всей вселенной, указанное спектрометрическими исследованиями, так как по существу установится обмен — чрез посредство эфира — между всеми мирами. Исследование упругости или сжимаемости газов под малыми давлениями, задуманное мною в 70-х годах и отчасти тогда же выполненное, имело, между прочим, целью проследить, насколько то возможно для имеющихся способов измерений малых давлений, изменения в газах, находящихся под малыми давлениями. Подмеченные для всех газов (мною с М. Л. Кирпичевым, 1874) так называемые положительные отступления от Бойль—Мариоттова закона, затем подтвержденные многими и, между прочим, Рамзаем (хотя до сих пор и непризнаваемые еще некоторыми исследователями), до некоторой степени указывают на единообразие поведения всех газов и на стремление их при уменьшении давления к некоторому пределу в расширении, как есть предел для сгущения — в сжижении и критическом состоянии.[4] Но в наблюдении очень малых давлений встретились непреоборимые трудности, тем большие, что для определения очень малых давлений оказалось невозможным заменить ртуть более легкими жидкостями (например, серною кислотою или нефтяными маслами), потому что они оказались способными выделять из себя в манометрическую пустоту ничтожно малые, однако ясно видимые количества каких-то газов, хотя были предварительно неделями выдержаны при 100° в пустоте, доставляемой лучшими насосами. Таким образом практически оказалось невозможным сколько-либо точно измерять давления, меньшие, чем в десятые доли миллиметра высоты ртутного столба, а это — когда дело идет о разрежениях, подобных тем, какие надо предполагать даже на высоте 50 километров над уровнем наших морей — чересчур большие величины. Поэтому представление об эфире как сильно-разреженном газе атмосферы, не может доныне подлежать опытному исследованию и измерению, которые одни способны наводить (индуцировать) мысль на правильные пути и приводить затем к следствиям, опять подлежащим опытной и измерительной поверке.
Но и помимо этого, представление о мировом эфире, как предельном разрежении паров и газов, не выдерживает даже первых приступов вдумчивости — в силу того, что эфир нельзя представить иначе, как веществом, все и всюду проникающим; парам же и газам это не свойственно. Они сгущаемы при увеличении давлений, и их нельзя представить содержащимися во всех веществах, хотя они и широко распространены во всех телах природы, даже в аэролитах. Притом — и это, всего важнее — они, по своей химической природе и по своим отношениям к другим веществам, бесконечно разнообразны; эфир же однообразен всюду, насколько то нам известно. Будучи разнородны по своим химическим свойствам, известные нам пары и газы должны были бы химически разнообразно воздействовать на тела, которые они проникают, если бы эфир был их совокупностью.
Прежде чем итти далее, считаю неизбежно необходимым оговориться в отношении здесь и далее вводимых мною химических соображений. Избежать их при обсуждении мирового эфира было трудно, но во времена Галилея и Ньютона еще возможно. Ныне же это было бы противно самым основным началам дисциплины естественной философии, потому что со времен Лавуазье, Дальтона и Авогадро—Жерара химия получила все высшие права гражданства в обществе наук о природе и, поставив массу (вес) вещества во главе всех своих обобщений, пошла за Галилеем и Ньютоном. Мало того, чрез химию, только при ее приемах, действительно вкоренилось во всем естествознании стремление искать решение всяких задач, касающихся конечных, измеримых тел и явлений, в постижении взаимодействия беспредельно малых их отдельностей, называемых атомами, но в сущности (по реальному представлению) мыслимых, как химически неделимые индивидуумы, ничего общего не имеющих с механически-неделимыми атомами древних метафизиков. Доказательства этому последнему многочисленны, но достаточно упомянуть о том, что современные атомы не раз объясняли вихревыми кольцами (vortex), что и поныне живо стремление понять сложение химических атомов или друг из друга, или из «первичной материи» и что как раз в последнее время, особенно по поводу радиоактивных веществ, стали признавать деление химических атомов на более мелкие «электроны», а все это логически не было бы возможно, если бы «атомы» признавались механически неделимыми. Химическое миросозерцание можно выразить образно, уподобляя атомы химиков небесным телам: звездам, солнцу, планетам, спутникам, кометам и т. п. Как из этих отдельностей (индивидуумов) слагаются системы, подобные солнечной или системам двойных звёзд, или некоторым созвездиям (туманностям) и т. п., так представляется сложение из атомов целых частиц, а из частиц тел и веществ. Это для современной химии не простая игра слов или не одно уподобление, а сама реальность, руководящая всеми исследованиями, всякими анализами и синтезами химии. У нее свой микрокосм в невидимых областях, и, будучи архиреальною наукою, она все время оперирует с невидимыми своими отдельностями, вовсе не думая считать их механически неделимыми. Атомы и частицы (молекулы), о которых неизбежно говорится во всех частях современной механики и физики, не могут быть чем-либо иным, как атомами и частицами, определяемыми химией, потому что того требует единство познания. Поэтому и метафизика нашего времени, если желает помогать познанию, должна понимать атомы так же, как их понимать могут естествоиспытатели, а не на манер древних метафизиков китайско-греческого образца. Если Ньютоново всемирное тяготение реально раскрыло силы, всегда действующие даже на беспредельно больших расстояниях, то познание химии, внушенное Лавуазье, Дальтоном и Авогадро—Жераром, раскрыло силы, всегда действующие на неизмеримо малых расстояниях, и показало как громадность этих сил (что видно, например, из того, что силами этими легко сжижаются газы, подобные водороду, едва недавно сжиженному совокупностью физических и механических усилий), так и превращаемость их во все прочие виды проявления энергии, так как химическими силами (например при горении) достигаются механические и физические. Поэтому все современные основные понятия естествознания — следовательно, и мировой эфир — неизбежно необходимо обсудить под совокупным воздействием сведений механики, физики и химии, и, хотя понятие об эфире родилось в физике, и хотя скептическая индифферентность старается во всем усмотреть «рабочую гипотезу», вдумчивому естествоиспытателю, ищущему саму действительность, какова она есть, и не довольствующемуся смутными картинами волшебного фонаря фантазии, хотя бы украшенного логичнейшим анализом, нельзя не задаваться вопросом: что же такое это за вещество в химическом смысле?
Моя попытка и начинается с этого вопроса.
Ранее, чем излагать свой посильный ответ на вопрос о химической природе эфира, считаю долгом высказаться о мнении, которое читал между строк и не раз слыхал от своих учёных друзей, верящих в единство вещества химических элементов (или простых тел) и в происхождение их из одной первичной материи. Для них эфир содержит эту первичную материю в несложившемся виде, то есть не в форме элементарных химических атомов и образуемых ими частиц и веществ, а в виде составного начала, из которого сложились сами химические атомы. Нельзя не признать в таком воззрении увлекательной стороны. Как миры представляют иногда сложившимися из разъединенных тел (твердой космической пыли, болидов и т. п.), так атомы представляют происшедшими из первичного вещества. Сложившиеся миры остаются, но рядом с ними остается в пространстве космическая пыль, кометы, болиды и т. п. материалы, из которых предполагается их сложение уже многими. Так остаются и сложившиеся атомы, но рядом с ними сохранился и между ними движется их материал, то есть всепроникающий и первозданный эфир. Одни при этом полагают, что есть ряд видимых явлений, при которых атомы рассыпаются в свою пыль, то есть в первичную материю, как рассыпаются кометы в потоки падающих звезд. Химики и физики, так думающие, представляют, что как геологические изменения или как сложение и распадение миров идут перед нашими глазами, так пред нами же в тиши разрушаются и вновь слагаются и атомы в своей вечной эволюции. Другие, не отрицая такой возможности — в виде особо редкого и исключительного случая, считают мир атомов сложенным в твердь прочно и полагают невозможным направить опыт на то, чтобы уловить это, то есть считают невозможным на опыте рассыпать атомы в первичную материю или образовать из неё на наших глазах новые атомы химических элементов, то есть процесс их происхождения понимают раз бывшим и законченным навсегда, а в эфире видят остатки, отбросы. С последними — реалистами не приходится считаться, потому что при таком представлении мыслители руководятся не следствиями из наблюдений или опытов, а только воображением, свобода которого обеспечена в республике науки. Но с первыми, то есть с истинными поклонниками продолжающейся эволюции вещества атомов, считаться химическому реализму неизбежно, потому что исходные положения нашей науки состоят не только в том, что вся общая масса вещества постоянна, но постоянны и те формы вещества, которые понимаются как элементарные атомы и в отдельности являются как «тела простые», признаваемые неспособными превращаться друг в друга. Если бы эфир происходил из атомов и атомы из него слагались, то нельзя было бы отрицать образование новых, небывалых атомов и должно было бы признавать возможность исчезания части простых тел, взятых в дело, при тех или иных наблюдениях и опытах. Давно- давно масса людей, по старому предрассудку, верит в такую возможность и, если бы это мнение не сохранялось в наши дни, не являлись бы Емменсы в С. А. С. Штатах, стремящиеся, по манере алхимиков, превратить серебро в золото, или такие учёные, как Фиттика (F. Fittica), в Германии, который еще недавно, в 1900 году, старался доказывать, что фосфор может превращаться в мышьяк. Множество случаев подобного превращения одних простых тел в другие описывалось в те 50 лет, в течение которых я внимательно слежу за химической литературой. Но каждый раз, при тщательном исследовании подобных случаев, оказывалась или простая ошибка предубеждения, или недостаточная точность исследования, и вновь[5] защищать индивидуальную самобытность химических элементов я здесь не предполагаю. Мне следовало, однако, напомнить об этом, рассматривая эфир, потому что, помимо химической бездоказательности, мне кажется, невозможно сколько-либо реальное понимание эфира, как первичного вещества, потому что у веществ первейшими принадлежностями должно считать массу или вес и химические отношения: — первую для понимания большинства явлений при всех расстояниях, вплоть до бесконечно больших, а вторые — при расстоянии неизмеримо малых или соизмеримых с величинами тех мельчайших отдельностей, которые называют атомами. Если бы дело шло об одном том эфире, который наполняет пространство между мировыми телами (солнцем, планетами и т. п). и передает между ними энергию, то можно было бы — с грехом пополам, ограничиваться только предположением о массе, не касаясь его химизма, можно было бы даже считать эфир содержащим «первичную материю», как можно говорить о массе планеты, не касаясь ее химических составных начал. Но вполне, так сказать, бескровный, ближе ничем не определяемый эфир окончательно теряет всякую реальность и составляет причину беспокойства вдумчивых естествоиспытателей, лишь только спускаемся с неба на землю и признаем его проникающим все тела природы. Необходимость лёгкого и полного проникновения всех тел эфиром следует признать не только ради возможности понимания множества общеизвестных физических явлений, начиная с оптических (над чем не считаю надобным останавливаться), но и по причине великой упругости и, так сказать, тонкости эфирного вещества, атомы которого всегда и все представляют себе не иначе, как очень малыми сравнительно с атомами и частицами химически известных веществ, то есть подобными аэролитам среди планет. Притом такая проницаемость эфиром всех тел объясняет и невозможность уединить это вещество, как нельзя собрать ни воды, ни воздуха в решете, каким для эфира должно считать всякие твердые или иные вещества и преграды. Способность эфира проникать всюду, во все тела можно, однако, понимать, как высшую степень развития того проникновения газов чрез сплошные преграды, которое Грем изучал для каучука в отношении многих газов, а Девилль и др. нашли для железа и платины по отношению к водороду[6].
Обладая малым весом атома и низшею из всех известных газов плотностью, водород не только вытекает или диффундирует сильнее или быстрее всяких других газов чрез малейшие отверстия, но способен проникать и чрез сплошные стенки таких металлов, как платина и особенно палладий, чрез которые другие газы не проникают. Но тут несомненно действует не только быстрота движения частиц водорода, тесно связанная с его малою плотностью, но и химическая способность того же разряда, которая проявляется как при образовании сложных тел, содержащих водород, так и при образовании растворов, сплавов и тому подобных, так называемых, неопределенных соединений. Механизм этого проникновения можно представить подобным — на поверхности проницаемого тела — растворению газа в жидкости, то есть вскакиванию его частиц в промежутки между частицами жидкости, замедлению движения (отчасти некоторому сгущению газа) и такому или иному согласованию движений обоих видов частиц. В массе проницаемого тела сжатый газ, поглощенный на поверхности прикосновения, конечно, распространяется во все стороны, диффундируя от слоя к слою, если в опытах Робертс-Аустена даже золото диффундировало в твердом свинце на основании тех же сил. Наконец, на другой поверхности проницаемого тела сжатый газ находит возможность вырваться на большую свободу и, пока будет накопляться до исходного давления, станет проникать туда, где его нет или где его мало, то есть входить в преграду будет более со стороны превышающего давления, чем в обратном направлении. Когда же давления уравняются, наступит не покой, а подвижное равновесие, то есть с каждой стороны в преграду будет проникать и выбывать одинаковое число частиц или атомов. Допуская, а это необходимо, проницаемость эфира в отношении ко всем веществам, должно приписать ему, прежде всего, лёгкость и упругость, то есть быстроту собственного движения, еще в большем развитии, чем для водорода, и, что всегда важнее, ему должно приписать еще меньшую, чем для водорода, способность образовать с проницаемыми телами определённые химические соединения, так как эти последние характеризуются именно тем, что разнородные атомы образуют системы или частицы, в которых вместе или согласно движутся различные элементы, как солнечная система характеризуется зависимым, согласным и совместным движением образующих ее многих светил. А так как надо предполагать, что такое совместное движение водорода, например, с палладием, им проницаемым, действительно совершается для тех атомов водорода, которые находятся в среде атомов палладия, и что водород с палладием дает своё определённое соединение Pd²H (или какое иное), но при нагревании оно легко диссоциирует, то следует, мне кажется, допустить, что атомы эфира в такой высокой мере лишены этой, уже для водорода слабой, способности к образованию определённых соединений, что для них всякая температура есть диссоциационная, а потому ничего, кроме некоторого сгущения в среде атомов обычного вещества, для эфира признать нельзя.
Такое допущение, то есть отрицание для вещества или для атомов эфира всякой склонности к образованию сколько-либо стойких соединений с другими химическими элементами, ещё несколько лет тому назад должно было бы считать совершенно произвольным, а потому и мало вероятным даже гипотетически, так как все известные еще недавно простые тела и элементы, так или иначе, труднее или легче и прочнее или шатче, прямо или косвенно вступали во взаимные соединения, и тогда представить вещество, вовсе лишенное склонности подвергнуться под влиянием других веществ каким-либо химическим изменениям и чуждое способности образовать сложные частицы, — было бы чересчур смело и лишено всякой реальности, то есть чуждо известной действительности. Но вот в 1894 г. лорд Релей и проф. Рамзай открывают в воздухе аргон и определяют его, как недеятельнейшее из всех известных газообразных и всяких иных веществ. Скоро затем последовало открытие Рамзаем гелия, который по его яркому спектру Локьер предчувствовал, как особое простое тело на солнце; а затем Рамзай и Траверс открыли в сжиженном воздухе еще три таких же недеятельных, как аргон, газа: неон, криптон и ксенон, хотя содержание их в воздухе ничтожно мало и должно считаться для гелия и ксенона миллионными долями по объему и весу воздуха[7]. Для этих пяти новых газов, составляющих, вместе с открытием радиоактивных веществ, одни из блистательнейших опытных открытий конца XIX века, до сих пор не получено никаких сложных соединений, хотя в них ясно развита способность сжижаться и растворяться, то есть образовать так называемые неопределённые, столь легко диссоциирующие, соединения. Поэтому ныне, с реальной точки зрения, уже смело можно признавать вещество эфира лишённым — при способности проникать все вещества — способности образовать с обычными химическими атомами какие-либо стойкие химические соединения. Следовательно, мировой эфир можно представить, подобно гелию и аргону, газом, не способным к химическим соединениям.
Оставаясь на чисто химической почве, мы старались сперва показать невозможность понимания эфира ни как рассеянный пар или газ всюду распространенных веществ, ни как атомную пыль первичного вещества, из которого нередко еще доныне многие при знают сложение элементарных атомов, а потом пришли к заключению о том, что в эфире должно видеть вещество, лишённое способности вступать в сколько-либо прочные определенные химические соединения, что свойственно недавно открытым гелию, аргону и их аналогам.
Это первый этап на нашем пути; на нем, хотя недолго, необходимо остановиться. Когда мы признаем эфир газом — это значит прежде всего, что мы стремимся отнести понятие о нём к обычным, реальным понятиям о трёх состояниях веществ: газообразном, жидком и твердом. Тут не надо признавать, как то делает Крукс, особого четвёртого состояния, ускользающего от реального понимания природы вещей. Таинственная, почти спиритическая подкладка с эфира при этом допущении скидывается. Говоря, что это есть газ, очевидно, мы признаём его «жидкостью» в широком смысле этого слова, так как газы вообще суть упругие жидкости, лишённые сцепления, то есть той способности настоящих жидкостей, которая проявляется в виде свойства образовать — в силу сцепления — капли, подниматься в волосных (капиллярных) трубках и т. п. У жидкостей мера сцепления есть определённая, конечная величина, у газов она близка к нулю или, если угодно, величина очень малая. Если эфир — газ, то, значит, он имеет свой вес; это неизбежно приписать ему, если не отвергать ради него всей концепции естествознания, ведущего начало от Галилея, Ньютона и Лавуазье. Но если эфир обладает столь сильно развитою проницаемостью, что проходит чрез всякие оболочки, то нельзя и думать о том, чтобы прямо из опыта найти его массу в данном количестве других тел, или вес его определённого объема — при данных условиях, а потому должно говорить не об невесомом эфире, а только о невозможности его взвешивания. Конечно, тут скрыта своя гипотеза, но совершенно реальная, а не какая-то мистическая, внушающая сильное беспокойство вдумчивым естествоиспытателям.
Всё предшествующее, мне кажется, не только не противоречит общераспространённому представлению о мировом эфире, но прямо с ним согласуется. Добавка, нами сделанная, стремящаяся ближе реализовать понятие об эфире, состоит только в том, что мы пришли к необходимости и возможности приписать эфиру свойства газов, подобных гелию и аргону, и в наивысшей мере неспособность вступать в настоящие химические соединения. Над этим понятием, составляющим центральную посылку моей попытки, необходимо остановиться подробнее, чем над какою-либо иною стороною сложного и важного предмета, например, над сопротивлением эфирной среды движению небесных светил, над следованием за Бойль—Мариоттовым или Ван-дер-Ваальсовым законом, над громадною упругостью массы эфира, над мерою его сгущения и упругостью в разных телах и в небесном пространстве и т. п. Все такие вопросы придётся так или иначе умственно решать и при всяком ином представлении об эфире, как весомом, но не взвешиваемом веществе. Мне кажутся все эти стороны доступными для реального обсуждения уже ныне, но они завлекли бы нас слишком далеко и все же основной вопрос — о химическом составе эфира — остался бы при этом висеть в пустоте, а без него не может быть, на мой взгляд, никакой реальности в суждении об эфире; после же такого или иного ответа на этот вопрос, быть может, удастся двинуться дальше в реальном понимании других отношений эфира. Поэтому далее я стану говорить только о своей попытке понять химизм эфира, исходя из двух основных положений, а именно: 1) эфир есть легчайший — в этом отношении предельный — газ, обладающий высокою степенью проницаемости, что в физико-химическом смысле значит, что его частицы имеют относительно малый вес и обладают высшею, чем для каких-либо иных газов, скоростью своего поступательного движения, [8] и 2) эфир есть простое тело, лишённое способности сжижаться и вступать в частичное химическое соединение и реагирование с какими-либо другими простыми или сложными веществами, хотя способное их проницать, подобно тому, как гелий, аргон и их аналоги способны растворяться в воде и других жидкостях.
Дальнейшие стороны моей попытки — понять природу эфира — так тесно связаны с гелием, аргоном и их аналогами и с периодическою системою элементов, что мне ранее, чем итти вперед, необходимо особо остановиться над этими предметами и их взаимною связью.
Когда, в 1869 г., на основании сближений, подмеченных уж Дюма, Ленсеном, Петтенкофером и другими, между величинами атомных весов сходственных элементов, мною была выставлена периодическая зависимость между свойствами всех элементов и их истинными (то есть по системе Авогадро—Жерара с дополнениями Канницаро и с изменениями, вызываемыми периодическою законностью) атомными весами, не только не было известно ни одного элемента, неспособного образовать определенные сложные соединения, но нельзя было даже и подозревать возможности существования подобных элементов. Поэтому в периодической системе, данной мною в том виде, какой она сохранила и до сих пор, а именно при расположении по группам, рядам и периодам (см. 1-е издание книги моей «Основы химии», выпуск 3-й, вышедший в 1870 году, и статьи мои в журнале Русского Химического Общества 1869 г.), система элементов начиналась с группы 1-й и с ряда 1-го, где помещался и до сих пор помещается водород, легчайший из элементов, судя по атомному весу, и легчайший газ, судя по плотности, — при данных давлении и температуре. Никогда мне в голову не приходило, что именно водородом должен начинаться ряд элементов, хотя легче его не было и еще поныне между известными нет ни одного другого элементарного или сложного газа. Оставаясь на реальной почве, я решался предсказывать не только существование неизвестных элементов в среде известных, но и их свойства, как химические, так и физические, для них самих в свободном состоянии (простых тел) и для их соединений. Это, как известно, оправдалось последующими открытиями: галлия — Лекоком де Боабодраном, скандия — Нильсоном и, блистательнее всего, германия — Клементом Винклером, моим (ныне уже скончавшимся) хорошим другом и научным собратом. Предсказания эти были, по существу, тем, что называется в математике интерполированием, то есть нахождением, промежуточных точек на основании крайних, когда известен закон (или направление кривой, его выражающей), по которому точки следуют друг за другом. Поэтому оправдание предсказанного есть не что иное, как способ утверждения законности, и, следовательно, теперь можно смело полагаться на то, что в 1869—1871 гг. было только вероятным, и уверенно признавать, что химические элементы и их соединения находятся в периодической зависимости от атомных весов элементов. Эксполировать, то есть находить точки вне пределов известного, нельзя было на основании еще неупроченной законности. Но когда она утвердилась, можно на это решиться, и то, что дальше будет сказано об эфире, как элементе, гораздо более лёгком, чем водород, составляет такое эксполирование. Решимость моя, при той осторожности, какая должна быть свойственна всякому деятелю науки, определяется двумя соображениями. Во-первых, я думаю, что откладывать — по старости лет — мне уже нельзя. А, во-вторых, за последнее время стали много и часто говорить о раздроблении атомов на более мелкие электроны, а мне кажется, что такое дробление должно считать не столько метафизическим, сколько метахимическим представлением, вытекающим из отсутствия каких-либо определенных соображений, касающихся химизма эфира, и мне захотелось на место каких-то смутных идей поставить более реальное представление о химической природе эфира, так как, пока что-нибудь не покажет либо превращения обычного вещества в эфир и обратно, либо превращения одного элемента в другой, всякое представление о дроблении атомов должно считать, по моему мнению, противоречащим современной научной дисциплине, а те явления, в которых признаётся дробление атомов, могут быть понимаемы, как выделение атомов эфира, всюду проникающего и признаваемого всеми. Словом, мне кажется, хотя рискованным, но своевременным говорить о химической природе эфира, тем более, что, сколько мне известно, об этом предмете еще никто не говорил более или менее определённо. Когда я прилагал периодический закон к аналогам бора, алюминия и кремния, я был на 33 года моложе, во мне жила полная уверенность, что рано или поздно предвидимое должно непременно оправдаться, потому что мне все там было ясно видно. Оправдание пришло скорее, чем я мог надеяться. Теперь же у меня нет ни прежней ясности, ни бывшей уверенности. Тогда я не рисковал, теперь рискую. На это надобна решимость. Она пришла, когда я видел радиоактивные явления, как объяснено в конце статьи, и когда я сознал, что откладывать мне уже невозможно и что, быть может, мои несовершенные мысли наведут кого-нибудь на путь более верный, чем тот возможный, какой представляется моему слабеющему зрению.
Первоначально я выскажусь о положении гелия, аргона и их аналогов в периодической системе элементов, потом о представляемом мною месте эфира в той же системе, а закончу несколькими беглыми замечаниями по поводу ожидаемых свойств эфира, основанных на понятии о нём, выводимом из его положения в этой системе.
Когда в 1895 г. дошли до меня первые сведения об аргоне и его беспримерной химической инертности (он ни с чем, ни при каких условиях не реагирует), мне казалось законным сомневаться в элементарной простоте этого газа, и я предполагал, что аргон можно считать полимером азота N³, как озон О³ есть полимер кислорода О² , но с тем различием, что озон происходит, как известно, из кислорода с присоединением — как говорится — тепла, то есть выделяет на данный свой вес более тепла, вступая в реакции, одинаковые с кислородом, чем кислород при том же весе, а аргон можно было представить, как азот, потерявший тепло, то есть еще менее энергичный, чем обычный азот. Этот последний всегда служил в химии образцом химической инертности, то есть простым телом, очень трудно вступающим в реакции, и если бы представить, что его атомы, уплотняясь при полимеризации из N² в N³ , теряют теплоту, можно было ждать вещества еще в высшей мере инертного, то есть ещё более сопротивляющегося воздействию других веществ. Так, кремнезём, происходящий с отделением тепла из кремния и кислорода, менее последних способен к химическим реакциям. Подобное же представление о природе аргона и о связи его с азотом высказано было затем известнейшим ученым Вертело. Теперь, уже давно, я отказался от такого мнения о природе аргона и соглашаюсь с тем, что это есть самостоятельное элементарное вещество, как это с самого начала утверждал Рамзай. Поводов к такой перемене было очень много. Главнейшими служили: 1) несомненная уверенность в том, что плотность аргона гораздо менее 21, а именно, вероятно, лишь немногим более 19, если плотность водорода принять за 1, а для N³ надо ждать плотности около 21, так как вес частицы N³ =3·14=42, а плотность близка к половине веса частицы; 2) гелий, открытый тем же Рамзаем в 1895 г., представляет плотность, по водороду, около 2-х и обладает такою же полною химическою инертностью, как и аргон, а для него нельзя уже было реально мыслить о сложности частицы и ею объяснять инертность; 3) такую же инертность Рамзай и Траверс нашли для открытых ими неона, криптона и ксенона, и что пригодно было для аргона — было не применимо к ним; 4) самостоятельные особенности спектра каждого из указанных пяти газов, при полной их неизменности от ряда электрических искр, убеждали, что это целая семья элементарных газов, глубоко отличающихся от всех, до тех пор известных, своею полною химическою инертностью, и 5) постепенность и определенность физических свойств в зависимости от плотности и от веса атома[9] дополняют, благодаря трудам того же Рамзая, уверенность в том, что здесь дело идет о простых телах, самобытность которых, при отсутствии химических превращений, и можно было утверждать только постоянством физических признаков. Укажем для примера на изменение температуры кипения (при давлении в 760 миллим.) или той, при которой достигается упругость, равная атмосферной, и могут существовать — при указанном давлении — как жидкая, так и газообразная фазы:
Химич. знак и состав частицы Гелий.
He Не

Категория «Дизайнеры»
Взлеты Топ 5
|
| ||
|
+76 |
114 |
zo0m's journal : Немного меня... |
|
+73 |
99 |
Александр Никулин |
|
+72 |
104 |
Чеширская__кошка |
|
+43 |
117 |
Галерея web-дизайна Origin Site | RSS лента |
|
+4 |
5 |
TATARNIKOFF.COM |
Падения Топ 5
|
| ||
|
-3 |
120 |
Секреты мастераСекреты мастера |
|
-10 |
13 |
fashion-fashion.ru: всё самое модное |
|
-12 |
4 |
Дизайн и мебель |
|
-14 |
9 |
MisterM |
|
-15 |
8 |
PSDTUTS |
Популярные за сутки
Загрузка...
BlogRider.ru не имеет отношения к публикуемым в записях блогов материалам. Все записи
взяты из открытых общедоступных источников и являются собственностью их авторов.
взяты из открытых общедоступных источников и являются собственностью их авторов.